Die Zusammensetzung der Atmosphäre
Die Atmosphäre setzt sich aus
78% Stickstoss
20.95% Sauerstoff
0.93% Argon
0.04% CO2

Von diesen 0.04% werden durch die Natur 96% emittiert!
Das bedeutet, dass menschliche Emissionen 4% der 0.04% ausmachen. Das sind 0.0016% weltweit.
Der Schweizer Bundesrat und Grüne ungebildete, sowie vom Bundesrat, speziell von Frau Sommaruga (einer Klavierlehrerin) aus der Versenkung herbeigezauberte Pseudo Professoren welche die völlig lächerliche Behauptung vertreten, dass der Schweizer CO2-Anteil von ca. 2% der weltweiten 0.0016% einen Einfluss hätten, dabei misachten die die physikalische Gesetzmässigkeiten, die seit über 100 Jahren als gesichertes Wissen in den Universitäten gelehrt werden.
Was für ein Schwachsinn und vor allem der grösste Betrug der Schweizer Geschichte, verursacht oder absichtlich herbeigeführt durch komplett korrupte- oder unwissende Politiker und Ihre medialen Gehilfen, die sich seit langem als Hofnarren herabgelassen haben.
Warum lügen die Bundesräte, Parteien sowie Medien die Bevölkerung dermassen in einer Arroganz und völlig antiwissenschaftlichen Abbartigkeit an?
Ist es Korruption? Dummheit? Oder beides?
Diese bundesrätliche in der Scheizer Geschichte grösste Lüge kostet die Schweizer Haushalte pro Jahr über 4'000 CHF. Mit dem hohen Benzinpreis seit einigen Tagen haben sich diese Kosten auf CHF 6'000 erhöht. Und falls aus dieser äusserst dummen antiwissenschaftlichen Behauptungen heraus der politische Zwang entsteht, eine E-Schüssel zu kaufen, erhöhen sich die Kosten pro Haushalt auf 10'000 CHF! Ein nicht nur wirtschaftliches Desaster, sondern auch Umwelttechnisch und Energietechnisch, denn gleichzeitig reduziert man in ganz Europa die sicheren und kostengünstigen Energielieferanten KKWs. Ein weiterer politischer Schwachsinn, der die Schweizer Milliarden kostet und die Gefahr von Blackouts jeden Monat erhöhen lassen. Und die Entsorgung des Sondermülls, der aus den Abertonnen an Batteriemüll entsteht, ist nicht im geringsten gelöst. Die Reichweiten der E-Fahrzeuge beträgt im Winter bei 0 Grad noch genau 30% im Vergleich zu Benziner! Wer in die Ferien ins Ausland fahren will, sollte gleich mehrere Tage für das Laden der Akkus einplanen, denn eine direkte Fahrt ins Urlaubsziel wird niicht mehr möglich sein.
Man muss diese mit Abstand dümmste Politische Entwicklung aufhalten, sonst enden wir in einem Energie- und Wirtschaftsdesaster, welche die Schweiz noch nie erlebt hat!
Physikalische Eigenschaften des CO2 Moleküls
Wärmeaustausch
Beim Wärmeaustausch zwischen Atmosphäre und Erdoberfläche sind zwei grundsätzlich verschiedene Austauschmechanismen beteiligt.
Wärmeaustausch via Konvektion:
Die Wärme wird durch Stöße von Molekülen untereinander übertragen. Je schneller sich die Moleküle bewegen, desto mehr Energie, sprich Wärme kann übertragen werden, desto wärmer ist die Umgebungsluft. Die Art der Gasmoleküle ist dabei beliebig, d. h. Konvektionswärme kann von Stickstoff, Sauerstoff, Wasserdampf, Argon, Kohlendioxid, Methan, flüchtigen Kohlenwasserstoffen und anderen Gasen in der Atmosphäre übertragen werden. Konvektionswärme kann in der Gesamtbilanz nur von warm nach kalt fließen (2. Hauptsatz der Thermodynamik), da die Temperatur der Atmosphäre von unten nach oben abnimmt, kann keine Konvektionswärme aus der kalten Atmosphäre wieder zurück Richtung Erdoberfläche fließen und die Erdoberfläche erwärmen oder für eine verzögerte Wärmeabgabe verantwortlich sein. Dieser Mechanismus scheidet für eine Wärmeübertragung von der Atmosphäre in Richtung Erdoberfläche aus.
Wärmeaustausch via Infrarotstrahlung:
Infrarot (IR)-Strahlung ist eine elektromagnetische Strahlung, für deren Absorption und Emission bestimmte Voraussetzungen gelten. Ein Gasmolekül muss zwingend entweder ein permanentes oder ein sich während der Molekülschwingung veränderndes Dipolmoment aufweisen. Stickstoff, Sauerstoff und Argon haben weder ein permanentes noch ein sich während der Molekülschwingung veränderndes Dipolmoment, sie können daher Infrarotstrahlung weder absorbieren noch emittieren.
Bei der Absorption von Infrarotstrahlung geht das Molekül vom Grundzustand in einen angeregten Schwingungszustand über, bei der Emission von Infrarotstrahlung geht das Molekül vom angeregten Schwingungszustand in der Grundzustand zurück und die überschüssige Energie wird dabei als IR-Photon abgestrahlt.
Kohlendioxid ist ein Molekül aus einem Kohlenstoff- und zwei Sauerstoffatomen die linear und symmetrisch angeordnet sind. Es besitzt daher kein permanentes Dipolmoment. Da sich das Dipolmoment während einer asymmetrischen Schwingung verändert, kann Kohlendioxid Infrarotstrahlung absorbieren und sie wieder emittieren. Kohlendioxid besitzt ausschließlich zwei Absorptionsbanden, die von zwei energiegleichen asymmetrischen Schwingungen (2350 cm-1) und einer Beugeschwingung (670 cm-1) her rühren. Höher angeregte Zustände sind beim Kohlendioxid bei den atmosphärischen Temperaturen nicht besetzt.
Da alle Kohlendioxidmoleküle (analoges gilt für Wasser) gleich aufgebaut sind erhält man für alle Kohlendioxidmoleküle auch die gleichen Absorptions/Emissions Wellenlängen und nicht etwa ein kontinuierliches Spektrum der Infrarotstrahlung.
Aufgrund der von der Erdoberfläche abgestrahlten und der dann vom anthropogenen Kohlendioxid zurück gestrahlten Infrarotstrahlung soll es nach Meinung einiger Klimaforscher (z.B. im IPCC und im PIK) zu einer steigenden Erwärmung der Erdoberfläche kommen.
Jedoch kann ein Molekül Infrarotstrahlung nur bei der Wellenlänge emittieren, deren Wellenlänge es absorbieren kann (analog einem Rundfunksender und Empfänger deren Frequenzen ebenfalls aufeinander abgestimmt sein müssen. Liegen die Frequenzen auch nur minimal auseinander, dann ist kein Radioempfang möglich). Dieses grundlegende physikalisch-chemische Sender/Empfänger Prinzip stellt die Schlüsselrolle des Wärmeaustausches zwischen der Atmosphäre und der Erdoberfläche dar. Kohlendioxid in der Erdatmosphäre kann also nicht eine x-beliebige Wärmestrahlung, die von der Erdoberfläche abgestrahlt wird absorbieren und wieder Richtung Erde emittieren und umgekehrt kann nicht jedes Molekül auf der Erdoberfläche von der durch Kohlendioxid emittierten Infrarotstrahlung erwärmt werden, weil es diese aufgrund seiner ihm eigenen Moleküleigenschaften gar nicht absorbieren kann. Trifft also das IR-Photon des Kohlendioxids auf einen Stoff, der bei dieser Frequenz nicht absorbiert, kann dieser Stoff auch nicht durch Kohlendioxid erwärmt werden.
Kohlendioxid absorbiert und emittiert bei 2350 cm-1 und 670 cm-1 und Wasser bei 3400 cm-1, 1650 cm-1 und 730 cm-1. Dies bedeutet nichts anderes, als dass Wasser und Kohlendioxid nur in einem sehr schmalen Bereich Wärmestrahlung austauschen können. Insbesondere führen die starken Absorptionen bei 2350 cm-1 (Kohlendioxid) und 3800 cm-1/1650 cm-1 (Wasser) zu keinerlei Wärmeaustausch zwischen diesen beiden Medien und es gibt somit kein Wärme-Pingpong zwischen diesen beiden Akteuren, das zu einer verzögerten Wärmeabgabe führen würde, wie dies oft behauptet wird. Oder einfach gesagt, das Kohlendioxid in der Atmosphäre lässt den Ozean völlig kalt.
Bei 670 cm-1 überlappen die Absorptionen von Wasser und Kohlendioxid zwar, ein Hin und Her an Wärme wäre denkbar. Allerdings ist sehr viel mehr Wasser in der Atmosphäre (ca. 50-100 Mal mehr) als Kohlendioxid. Um hier noch den Beitrag des Kohlendioxids heraus zurechnen muss man also äußerst exakt messen. Und selbst wenn es gelänge, was würde man mit einem Ergebnis anfangen, bei dem der Kohlendioxidbeitrag nur noch die alleroberste Spitze des i-Tüpfelchens darstellt? Es ist also nicht so, dass ausgerechnet das anthropogene Kohlendioxid quasi eine Lücke verstopft und die Abstrahlung von Wärme verhindert.
Hingegen wäre natürlich der Wärmeaustausch via Infrarotstrahlung zwischen Wassertropfen in der Atmosphäre und Wasser im Ozean denkbar, denn hier passen die Wellenlängen bei allen Absorptionsbändern exakt aufeinander. Wenn die vom Wasserdampf der Atmosphäre zurück gestrahlten IR-Photonen vom Wasser im Ozean wieder absorbiert werden, wird Wärme in den Ozean zurück transportiert. Diese Wärme wurde jedoch zuvor beim Abstrahlen des IR-Photons aus dem Ozean entnommen, so dass sich in der Gesamtbilanz keine Erwärmung ergeben kann! Weiterhin wird Infrarotstrahlung radial, das heißt in alle Richtungen abgestrahlt (und nicht etwa nur in Richtung der Erdoberfläche) die Hälfte der IR-Photonen verschwindet also auf Nimmerwiedersehen ins Weltall, was zu einer Abkühlung führt.
Hingegen haben das Edelgas Argon sowie Stickstoff und Sauerstoff nicht die Fähigkeit Wärme in das Weltall abzustrahlen.
Kohlendioxid ist also aufgrund seiner physikalisch-chemischen Eigenschaften tatsächlich nicht in der Lage, die Erde durch ein Hin und Her der Wärmestrahlung zu erwärmen, wie es von manchen sogenannten Klimaexperten gebetsmühlenartig behauptet wird. Solange es auf der Erde keine oder nur geringfügig Medien gibt, die bei Wellenlängen von 2350 cm-1 oder 670 cm-1 (Kohlendioxid) IR-Photonen absorbieren, spielt es überhaupt keine Rolle wie hoch die CO2-Konzentrationen in der Atmosphäre sind; sie sind nicht im Stande, die Erdoberfläche zu erwärmen. Es ist doch sehr auffällig, dass in der Politik oder von sogenannten Aktivisten immer nur behauptet wird, die Erwärmung würde am anthropogenen Kohlendioxidausstoß liegen, aber nie jemand auch nur im Ansatz die physikalisch-chemischen Vorgänge dazu erläutert.
Kohlendioxid besitzt eine hohe Dichte und wiegt etwa das 1,5fache von Luft. Damit ist es das schwerste Spurengas in der Luft. CO2 absorbiert in den Bereichen 2.75, 4.25 und um 15 Mikrometer wärme. Die aufnehmbare Wärme ist jedoch begrenzt, bzw. CO2 ist bereits mit der geringen Menge in Bodennähe in der Atmosphäre zu 90% mit Wärme gesättigt. Da CO2 schwerer als Luft ist, verhält es sich entsprechend und fliesst in der Regel in Richtung Bodennähe. Durch Konvektion wird CO2 in der untersten Atmosphärenschicht durcheinander gewirbelt. In ca. 10-12 Kilometer Höhe beträgt der Anteil an CO2 noch ca. 4-16 Moleküle auf 1 Million. Das Spurengas CO2 ist nicht nur für den Calvin-Zyklus elementar wichtig, sondern spielt auch für die Wärmeableitung in der Atmosphäre eine sehr wichtige Rolle. Umso mehr CO2 in höheren Luftschichten vorhanden ist, umso besser wird Wärme ins All abgeleitet. Getreu des 2. Thermodynamischen Hauptsatzes. Ausserdem wird CO2 für den Pflanzenwachstum zwingend benötigt. Ohne CO2 gäbe es auf der Erde keine Biosphäre. Umso mehr CO2 anteilig in der Atmosphäre vorkommt, desto grösser wird die Biosphäre. Dies hatten wir bereits im Artikel vom 17.11.2017 detailliert aufgezeigt und auf die Studie vom "Lead author Zaichun Zhu from Peking University" hingewiesen.
Eine Analyse der Physikers Peter Dierkes, an dieser Stelle nochmals vielen Dank für die Abhandlung
CO2-Anstieg in der Atmosphäre (Gegenwart): 1) durch Ausgasung aus dem Ozean / die VA (Verbrennung) kann die durch Photo-Synthese + BIO-Pumpe NICHT allein kompensiert werden; 2) es kann sein, dass sich das Upwelling (s.v.) in den letzten 100 a verstärkt hat (s. Änderungen der Wind-/Strömungsverhältnisse); 3) es könnte aber auch sein, dass CO2 aus dem Erdinneren austritt (s. es handelt sich um alten C / i.e. C-14 arm, Isotopenmethode); 4) Überblick-Kohlenstoffspeicher: Atmosphäre-2017, rd. 850 Gt C = 0,001% des globalen Gesamt-C (der CO2-Wert betrug rd. 406 ppmv) / Hydrosphäre-2017, rd. 38000 Gt C = 0,045% des globalen Gesamt-C / Lithosphäre-2017, rd. 90600 Gt C (anorganisch, organisch) / Biosphäre-2017, 😎 in Summe wird dieser CO2-Kreislauf/a auf 300-400 Gt CO2 geschätzt (kann durch die sog. ENSO-Aktivität = El Nino Southern Oscillation u.a. um rd. 10% schwanken); 9) El Nino tritt unregelmäßig auf (der Zentralpazifik wird dadurch wärmer = mehr CO2!) / s. 1998 gab es einen starken El Nino (die Global-T stieg um +0,6°C und die globale CO2-Menge um +3 ppm) / 1 ppm bedeutet rd. 8 Gt CO2; d.h. zusätzliche 24 Gt CO2 / s. dagegen 1992 hier gab es eine La Nina-Abkühlung (Pazifik), der CO2-Anstieg wurde auf +0,5 ppm reduziert (d.h. kaum ein Anstieg); 10) die menschlichen Emissionen spielen mit herein / diese liegen in der Größenordnung wie die ENSO-Schwankungen (s.o.) / der Anstieg war gleichmäßig von rd. 25 Gt (2000) auf 36 Gt (2014/15) / das entspricht einem Anstieg um 40%, wobei keine Auswirkung auf das Klima + globale T nachweisbar war / s. nach den Satellitenmessungen (RSS = Remote Sensing System, Datensysätze) wurde in den vergangenen 18 a kein ansteigender Trend bei der Global-T erkannt
Teil IX:
1) die Gesamtmenge von CO2 in der Atmosphäre nimmt zu (meist durch anthropogene Emissionen verursacht) / die Emissionen/a liegen nach NOAA (Daten am Mauna Loa, Hawaii) bei rd. 2 ppm und erreichten 2014 rd. 36 Gt CO2 (statistische Daten der Länder-Emissionen); 2.) allerdings blieben davon nur 2,3 ppm = 18,4 Gt in der Atmosphäre; 3) sollte sich das so fortsetzen, dann könnte sich der CO2-Gehalt in der Atmosphäre in 170-200 a verdoppeln (rd. 800 ppm); 4) nach der IPCC-Hypothese (fehlender Beweis) sollte eine Verdoppelung einen T-Anstieg um rd. 3°C (+/- 1,5°C) bewirken; 5) auch damit würde man in 2100 unter dem politischen "Grenzwert" von +2°C liegen ohne Reduktion der Emissionen (!); 6) der CO2-Anstieg (2x bis zu 200 a) ist nicht beunruhigend / teure politische Maßnahmen zur CO2-Reduktion sind nutzlos, da CO2 kaum-sehr geringen Klima-Effekt hat (s. das Heizwirkungs-Dilemma), sehr wohl aber positive Auswirkungen (s. Pflanzen, Nahrungsmittel etc.); 7) 1959: Franckes Lexikon der Physik, CO2 ist als Klima-Gas bedeutungslos; 8 ) die vergangenen 18 a mit insgesamt über rd. 500 Gt anthropogener CO2-Emissionen (ohne den vorhergesagten T-Anstieg) sind mit ein Beleg dafür.
Was ist CO2 und für was ist es zwingend notwendig?
Vorwort
Im Web liest man immer wieder, dass CO2 ein schädliches Klimagas sei. Wenn Sie diese Aussage irgendwo lesen, ist es keine wissenschaftlich korrekte Darstellung, sondern bloser Klima-Sektismus. CO2 ist und war zu keiner Zeit ein Klimagas sondern ein Spurengas und es ist auch in keinster Weise für irgendwas schädlich.
Die Atmosphäre besteht aus 78.1% Stickstoff, 20.9% Sauerstoff, 0.93% Argon und einige Spurengase. Beim Bildungsserver werden Kohlendioxid und Ozon, wie auf fast allen sogenannten Pseudo-Wissens-Portalen, als Treibhausgase eingestuft. Das ist schlichtweg falsch! Es handelt sich dabei um Spurengase, welche bereits durch das geringe Vorkommen in der Atmosphäre nicht in der Lage sind, dieser Behauptung gerecht zu werden.
CO2 kommt in der Atmosphäre mit 0.0403% vor. Der anthropogene Anteil davon ist ca. 4%. 4% von 0.0403% sind 0.001612%. Oft wird 400 ppm (0.04%) als Mengenmass angegeben. Das bedeutet nichts anderes als 400 Moleküle auf 1 Million Moleküle. Davon sind 16 CO2 Moleküle anthropogener Herkunft.
In höheren Luftschichten nimmt der Anteil des CO2 aufgrund seiner 1.5 mal schwereren Masse exponentiell ab und beträgt auf 10'000 Metern Höhe noch ca. 4 Moleküle auf 1 Million. Wobei auch die restlichen Luftmoleküle in dieser Höhe wesentlich geringer vorkommen, als in Bodennähe. Das bedeutet, dass die gleiche Anzahl Moleküle eine grössere räumliche Fläche einnehmen, als in Bodennähe. Grosse Lücken vermindern mechanische Kollisionen der Moleküle, so dass durch die Thermodynamik abgeleitete Strahlungswärme ungehindert entweichen kann. CO2 wirkt in grossen Höhen als zusätzlicher Ableitungsverstärker.
Im Infrarot-Bereich kann CO2 nur im 15 Micrometer Bereich Wärme absorbieren und emittiert diese wieder, wenn eine mechanische Kollision mit anderen Molekülen auftritt. Funktioniert jedoch nicht mit allen molekühlen der Atmosphäre. Es tritt deshalb eine Sättigung der Wärmeaufname auf, so dass CO2 praktisch keine Wärme mehr aufnimmt.
Online liest man praktisch nichts über Wasserdampf, ist jedoch mit 4 % eines der wirksamsten Klimagase. Natürlich wird dies fast nicht mehr erwähnt, es würde die Lüge der Klimakirche ja auffliegen lassen.
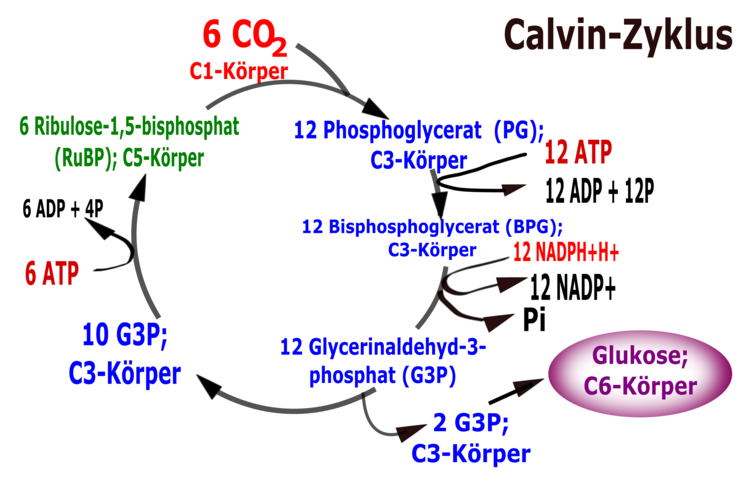
Calvin-Zyklus
Der nach dem Entdecker benannte CALVIN-Zyklus beschreibt innerhalb der Fotosynthese der Pflanzen den Weg des Kohlenstoffdioxids bis zur Entstehung eines Kohlenhydrats. Für den Ablauf der chemischen Reaktionen, die im Stroma des Chloroplasten stattfinden, werden als Voraussetzungen lediglich ATP als Energiequelle und NADPH + H+ als Reduktionsmittel benötigt. Licht ist für diesen Vorgang nicht nötig. Die komplexen Vorgänge werden in drei Phasen eingeteilt. Zunächst erfolgt die Fixierung des Kohlenstoffdioxids an einen Akzeptor (Ribulose-1,5-bisphosphat). Das erste daraus entstehende stabile Produkt ist ein Molekül mit drei Kohlenstoffatomen: Glycerinsäure-3-phosphat. Pflanzen, die auf diesem Weg Kohlenhydrate herstellen, nennt man daher C3-Pflanzen. Neben diesem Weg haben Fotosynthesespezialisten in Anpassung an trockene Umweltbedingungen Mechanismen entwickelt, Kohlenstoffdioxid in ihren Blattgeweben vorläufig zu konzentrieren.
In einer zweiten Phase erfolgt über Zwischenprodukte unter Verbrauch von ATP und mithilfe von NADPH + H+ die Reduktion der Glycerinsäure-3-phosphat zu Glycerinaldehyd-3-phosphat. Einige Moleküle dieses entstehenden Kohlenhydrats werden aus dem Kreisprozess ausgeschleust und sind die Grundlage für die Bildung weiterer Kohlenhydrate, Fette und Eiweiße u. a. zur Speicherung von Energie. Die verbleibenden Glycerinaldehyd-3-phosphat-Moleküle werden im Kreislauf zur Regeneration des Akzeptors unter nochmaligem Verbrauch von ATP verwendet.
Fotosynthetisch aktive Pflanzen zeigen auch einen Gaswechsel (Sauerstoffverbrauch und Kohlenstoffdioxidabgabe), der im Licht wesentlich aktiver abläuft als im Dunkeln und wegen seiner Ähnlichkeit zur Atmung als Lichtatmung bzw. Fotorespiration bezeichnet wird.
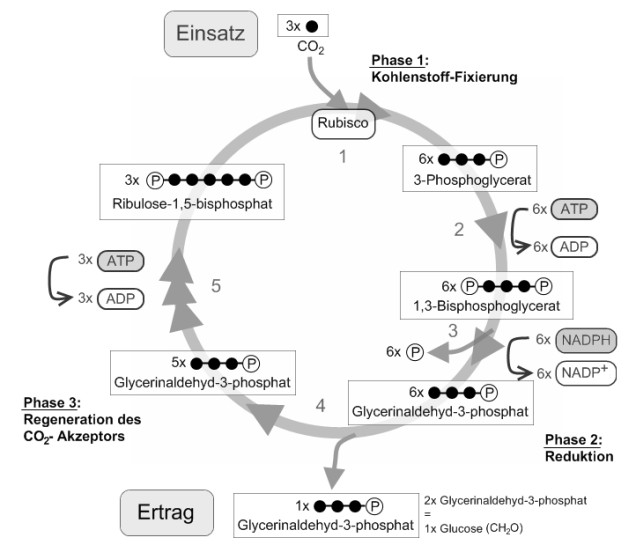
Ende der 40er-Jahre verfolgten CALVIN und seine Mitarbeiter, wie aus radioaktiv markiertem Kohlenstoffdioxid in einem Kreisprozess Kohlenhydrate hergestellt wurden. Der nach seinem Entdecker MELVIN CALVIN (1911-1997) benannte CALVIN-Zyklus, bzw. die lichtunabhängigen Reaktionen (Dunkelreaktion) der Fotosynthese, laufen im pigmentlosen Stroma des Chloroplasten ab. Als Voraussetzungen für die Dunkelreaktion dienen ATP als Energiequelle und NADPH + H+ als Reduktionsmittel, die in den lichtabhängigen Reaktionen (Lichtreaktion der Fotosynthese) bereitgestellt worden sind. Licht ist während dieses Prozesses nicht vonnöten.
Dabei ist zu beachten, dass sechs Kohlenstoffdioxidmoleküle in den Kreislauf eingeschleust werden müssen, um zwei Moleküle des Primärprodukts Glycerinaldehyd-3-phosphat für eine weitere Verwertung zu Glucose und anderen organischen Stoffen herzustellen.
Die ablaufenden chemischen Prozesse und die entstehenden Produkte sind dabei den Vorgängen der Zellatmung ähnlich.
Die komplexen Reaktionen des CALVIN-Zyklus werden in folgende Teilprozesse unterteilt:
Phase 1: Fixierung des Kohlenstoffdioxids (Carboxylierungsphase)
Den Einbau des Kohlenstoffdioxids in organische Verbindungen bezeichnet man als Kohlenstoffdioxidfixierung. Ribulose-1,5-bisphosphat, ein Zuckermolekül mit fünf Kohlenstoffatomen, das durch die Anlagerung zweier Phosphatgruppen besonders energiereich ist und als Akzeptor dient, bindet Kohlenstoffdioxid. Die dabei entstehende instabile Zwischenverbindung mit sechs Kohlenstoffatomen im Molekül zerfällt sofort in zwei Moleküle 3-Phosphoglycerinsäure. Dieses Molekül mit drei Kohlenstoff-Atomen ist das erste stabile Produkt im CALVIN-Zyklus.
Pflanzen, die nur auf diesem Weg Kohlenstoffdioxid binden können, werden zu den C3-Pflanzen zusammengefasst und sind die am häufigsten vertretenen unter den Pflanzen. Neben diesem Weg können Fotosynthesespezialisten (C4-Pflanzen, CAM-Pflanzen) in Anpassung an sonnige, trockene Standorte Kohlenstoffdioxid durch andere Mechanismen vorläufig als organische Säuren binden, bevor sie dann später mithilfe des CALVIN-Zyklus Kohlenhydrate daraus herstellen.
Die Bindung des Kohlenstoffdioxids an den Akzeptor
Ribulose-1,5-bisphosphat wird durch eine Carboxylase (Rubisco) gesteuert, die vermutlich weltweit mengenmäßig das häufigste Enzym ist.
Phase 2: Reduktion
Bei der Reduktion bindet die 3-Phosphoglycerinsäure eine bei der ATP-Spaltung frei werdende Phosphatgruppe. Die entstehende 1,3-Diphosphoglycerinsäure wird durch die Elektronen des NADPH + H+ zu Glycerinaldehyd-3-phosphat (G3P) reduziert, wobei aus der Carboxylgruppe (-COOH) die energiereichere Carbonylgruppe (-CHO) entsteht. Glycerinaldehyd-3-phosphat ist ein Kohlenhydrat mit drei Kohlenstoffatomen (Triose). Zwei Moleküle Glycerinaldehyd-3-phosphat werden aus dem Kreislauf zur weiteren Verwertung entlassen, wobei über mehrere Zwischenprodukte (z. B. Fructose-1,6-diphosphat) andere organische Stoffe sowie weitere Kohlenhydrate (Glucose, Saccharose), Fette und Eiweiße u. a. zur Speicherung von Energie entstehen.
Phase 3: Regeneration des Akzeptors
Während der Regeneration des Akzeptors entsteht aus den verbleibenden Glycerinaldehyd-3-phosphat-Molekülen zunächst über mehrere Zwischenprodukte, C3-,C4-, C5- und C7-Verbindungen, Ribulose-5-phosphat. Bevor dann daraus unter Spaltung von ATP und Anlagerung einer Phosphatgruppe der Akzeptor Ribulose-1,5-bisphosphat regeneriert wird.
Die lichtunabhängigen Reaktionen lassen sich für die Bildung eines Glucosemoleküls folgendermaßen zusammenfassen:
Die Coenzyme NADP+ und ADP + P werden durch die lichtabhängigen Reaktionen wieder regeneriert, sodass sie wieder für den Ablauf des CALVIN-Zyklus zur Verfügung stehen.
Wenn man alle Prozesse der lichtabhängigen und lichtunabhängigen Reaktionen berücksichtigt, ergeben sich folgende Gesamtgleichungen :
Dabei wird eine Energiemenge von 2 877 kJ benötigt, um ein Mol Glucose zu bilden.
Fotosynthetisch aktive Pflanzen zeigen einen zur Fotosynthese in Konkurrenz stehenden Prozess, der Lichtatmung oder Fotorespiration genannt wird. Ausgangspunkt für diesen Prozess ist die Doppelfunktion des Enzyms für die Fixierung von Kohlenstoffdioxid an den Akzeptor Ribulose-1,5-bisphosphat. Neben der Steuerung der Kohlenstoffdioxidfixierung ist es auch für die Bindung von Sauerstoff verantwortlich und wirkt als Oxygenase. Das Konzentrationsverhältnis von Sauerstoff- bzw. Kohlenstoffdioxid entscheidet über den Reaktionsverlauf.
Bei hoher Sauerstoff- aber geringer Kohlenstoffdioxidkonzentration wirkt das Enzym als Oxygenase und bindet Sauerstoff. Unter natürlichen Bedingungen (20 % , 0,03 % ) verlieren C3-Pflanzen ca. 50 % des assimilierten Kohlenstoffs, der energetisch nutzlos wieder in Kohlenstoffdioxid umgewandelt wird. Die Bedeutung dieses Prozesses ist bis heute unklar.
Bei geringem Sauerstoffgehalt im Vergleich zu hoher Kohlenstoffdioxidkonzentration überwiegt die Fixierung von Kohlenstoffdioxid, so wird die Fotorespiration gehemmt und die Nettofotosynthese steigt. Fotosynthesespezialisten (C4- und CAM-Pflanzen) zeigen im Gegensatz zu C3-Pflanzen keine Fotorespiration, da ihr Enzym für die Kohlenstofffixierung (PEP-Carboxylase) keinen Sauerstoff binden kann.
Kohlenstoffdioxid ist ein farb- und geruchloses Spurengas. Es ist mit einer Konzentration von ca. 0,04 % (derzeit 400 ppm entspr. 0,0403 %) ein natürlicher Bestandteil der Luft. Es entsteht sowohl bei der vollständigen Verbrennung von kohlenstoffhaltigen Substanzen unter ausreichendem Sauerstoff als auch im Organismus von Lebewesen als Kuppelprodukt der Zellatmung. Das CO2 wird dabei über den Atem abgegeben. Umgekehrt sind Pflanzen, manche Bakterien und Archaeen in der Lage, CO2 durch die Kohlenstoffdioxid-Fixierung in Biomasse umzuwandeln. So produzieren Pflanzen beispielsweise bei der Photosynthese aus anorganischem CO2 Glukose.
Biologische Aspekte des Spurengases CO2
Bei der Einatmung kommt Sauerstoff (O2) in die Lunge, bei der Ausatmung wird Kohlendioxid (CO2) aus der Lunge abgegeben. Im Inneren des Körpers geht die Atmung jedoch weiter, denn der Sauerstoff wird an die roten Blutkörperchen gebunden, wandert zu den Organen und wird dann gegen Kohlendioxid ausgetauscht. Dieses fließt dann mit dem Blut in die Lunge zurück, wo es abgeatmet wird. Da über das CO2 der Säure-Basen-Haushalt des Körpers mitgesteuert wird, wird in der Regel mit den Blutgasen gleichzeitig der Säure-Basen-Haushalt des Körpers mit erfasst. Hier spielen neben der Lunge auch die Niere und die Leber eine wichtige Rolle.
Kohlendioxid wird im Blut in drei Formen transportiert: im Plasma gelöst, als Bikarbonat, und an Proteine in Form von Carbaminoverbindungen gekoppelt.
Bicarbonat stellt den größten Anteil des CO2 im Blut dar (~88%).
Auf der venösen Seite der systemischen Kapillaren gelangt CO2 in die roten Blutkörperchen (RBC), wo es sich mit Wasser zu Kohlensäure (H2CO3) verbindet. Diese Reaktion wird durch das Enzym Carbonic Anhydrase (CA) katalysiert, das sich in RBC's befindet. Die Kohlensäure dissoziiert dann zu Bicarbonationen (HCO3-) und Wasserstoffionen (H+). Diese Reaktion tritt auch außerhalb der RBC's, im Plasma auf, aber sie ist viel langsamer aufgrund des Fehlens von CA.
CO2 + H2O ↔ H2CO3 ↔ H+ + HCO3-
In den Lungenkapillaren geschieht das Gegenteil: Bikarbonationen gelangen in den RBC und bilden zusammen mit H+ Kohlensäure, die in CO2 und H2O zerlegt wird, während CO2 in Alveolen diffundiert.
Zwei Faktoren machen den Bicarbonat-Puffer zum wichtigsten Puffersystem im Blut: Zum einen ist der Bikarbonatspiegel des Blutes ziemlich hoch (etwa 24 mmol/l), zum anderen ist es ein offenes System überschüssiges CO2 kann über die Lunge abgeatmet werden und der HCO3--Spiegel des Blutes kann durch die Niere reguliert werden. Auf diese Weise kann der pH-Wert des Blutes konstant gehalten werden (Säure-Basen-Homöostase).
Die Wichtigkeit des Bicarbonat-Puffers und seine körperweite Verbreitung wird durch Betrachtung des oxidativen Metabolismus des Körpers ersichtlich. Der oxidative Metabolismus produziert ständig CO2. Im Bicarbonat-Puffersystem wird dann CO2 mit H2O kombiniert, um Kohlensäure (H2CO3) zu bilden, die wiederum teilweise in H+-Ionen und Bikarbonat (HCO3-) zerfällt.
Obwohl die chemische Reaktion CO2 + H2O H2CO3 spontan stattfindet, wird sie auch durch das Enzym Carboanhydrase, eines der schnellsten bekannten Enzyme, erleichtert.
Der Bicarbonat-Puffer wirkt zusammen mit dem Ammonium-Puffer vor allem im Blutplasma. Intrazellulär herrschen andere Verhältnisse und dort sind vor allem der Phosphat-Puffer und der Protein-Puffer essentiell.
CO2 notwendig für Meeresbewohner
Die meisten Korallen brauchen zum Überleben sogenannte Zooxanthellen. Das sind Algen, die sich in der Außenhaut des Polypen ansiedeln. Alge und Polyp leben in einer Symbiose, also in einer biologischen Zweckgemeinschaft, die beiden Seiten Vorteile bringt. Die Zooxanthellen sind einfache, einzellige Lebensformen, die in einem direkten Stoffwechsel mit dem Polypen stehen. Mit Wasser und dem Kohlendioxid, das der Polyp ausscheidet, betreiben sie Photosynthese. Dabei nutzen sie die Energie der Sonnenstrahlen für einen chemischen Prozess, bei dem Sauerstoff und Glukose entstehen. Der Polyp braucht diese Stoffe zum Überleben.
Im Austausch erhalten die Zooxanthellen vom Polypen lebenswichtige Nährstoffe. Auf einer Außenfläche von einem Quadratzentimeter siedeln sich zirka eine Million dieser Algenzellen an. Sie sind es auch, die den Korallen ihre Farbe geben und deren Form bestimmen.
Da die Algen das Sonnenlicht für ihren Stoffwechsel brauchen, wächst die Koralle Richtung Sonne. Wenn eine im Riff unten gelegene Koralle in den Schatten einer weiter oben gelegenen kommt, verzweigt sie sich und wächst, wie die Äste eines Baums, Richtung Licht.
So kann es geschehen, dass genetisch identische Polypen gänzlich unterschiedlich aussehende Korallen bilden. Die Vielfalt der Anpassungsmöglichkeiten hat einen starken Einfluss auf ihr Aussehen.
"Wie bauen" die Korallen ein Riff?
Die Grundbausteine für sein Kalkskelett findet der Polyp im Meerwasser. Es sind Kalzium-Ionen und Kohlendioxid. Diese beiden Stoffe werden in den skelettaufbauenden Zellen des Polypen in Kalzium-Karbonat verwandelt. Je mehr Kohlendioxid die Zooxanthellen dem Stoffkreislauf der Koralle entnehmen, desto größer ist die Produktion von Kalzium-Karbonat.
Auch hier zeigt sich, wie gut die Symbiose zwischen Alge und Polyp funktioniert. Die Alge nutzt das Kohlendioxid, um durch die Photosynthese Nährstoffe für sich und den Polypen zu produzieren. Dabei hilft sie ihm gleichzeitig, sein Skelett auszubauen, das ihn und die Alge vor gefährlichen Umwelteinflüssen schützt.
Willis Eschenbach
Einer der Gründe, warum ich einen so großen Teil meines Lebens im tropischen Südpazifik verbracht habe, ist das Tauchen. Korallenriffe sind eines der erstaunlichsten Ökosysteme auf dem Planeten. Sie kochen über vor Energie, Bewegung und Farben. Ich habe hunderte und aberhunderte Stunden im Wasser verbracht, sowohl mit Schnorcheln als auch mit einem Atemgerät. Die meiste Zeit habe ich einfach nur die endlose Vielfalt der Riffbewohner bewundert, von kleinen und großen.

Infolgedessen habe ich ein langfristiges Interesse an den behaupteten Effekten gehabt, die Leute sagen, dass erhöhtes CO2 auf den Riffen hat. Durch die Zugabe von CO2 in die Luft wird das natürlich alkalische Meerwasser leicht neutralisiert. Beachten Sie, dass dies zwar eine Änderung des pH-Wertes durch die Erhöhung des CO2 ist, aber normalerweise als "Versauerung" des Ozeans bezeichnet wird, das ist nur eine alarmierende Terminologie. Da das zusätzliche CO2 den Ozean neutraler macht, kann es den Ozean nicht "versauern". Zumindest wenn die englische Sprache allgemein verstanden wird, kann etwas nicht neutraler und gleichzeitig "saurer" sein.]
In den letzten fünf Jahren oder so (siehe die Liste meiner früheren Beiträge in den letzten Anmerkungen), habe ich gesagt, dass die leichte Neutralisierung des Ozeans durch den anhaltenden Anstieg des CO2 für die Korallenriffe keinen Unterschied machen wird. Insbesondere habe ich festgestellt, dass sich der pH-Wert über Korallenriffen im Laufe einer Flut um einen vollen pH-Wert ändern kann. Ich habe auch darüber gesprochen, dass Korallenriffe oft eine CO2-Quelle sind und somit das Riff selbst den pH-Wert des Riffwassers herabsetzt (neutralisiert, fälschlicherweise als " säuerlich " bezeichnet). Meiner Meinung nach war es aufgrund dieser Tatsachen sehr unwahrscheinlich, dass eine kleine Neutralisierung des Ozeans einen signifikanten Unterschied in den Korallenriffen machen würde.
Da ich diese Trommel fünf Jahre lang geschlagen habe, war ich heute froh, einen Artikel auf Phys.org mit dem Titel Erhöhung des Säuregehalts zu sehen, der für Korallenriffe doch nicht schädlich sein könnte. Es wird ein gebührenpflichtiges Papier mit dem Titel Shifts in coral reef biogeochemistry and resulting acidification linked to offshore productivity diskutiert.
Der Phys.org-Artikel bringt es sehr deutlich zum Ausdruck:
Um besser zu verstehen, was mit Korallenriffen passieren könnte, wenn mehr Kohlendioxid durch einen Anstieg des Gases in der Atmosphäre durch menschliche Emissionen in die Ozeane gelangt, richteten die Forscher Überwachungsvorrichtungen entlang eines Korallenriffs vor der Küste der Bermudas ein - die Informationen der Sensoren wurden fünf Jahre lang (2007 bis 2012) beobachtet. Das Team hatte auch Zugriff auf Daten von einer Ozeanchemie-Überwachungsstation, die etwa 80 Kilometer von ihrem Studienort entfernt lag. Die kombinierten Daten boten eine einzigartige Perspektive auf die Korallenaktivität.
Bei der Untersuchung der Daten stellten die Forscher fest, dass es 2010 und 2011 zu Stacheln von Phytoplanktonblüten kam, die sich auf den Weg zum Korallenriff machten und mehr Nahrung als sonst für die Koralle boten. Die Koralle reagierte mit Wachstum, was dazu führte, dass sie mehr alkalisches Carbonat aus dem umgebenden Wasser zog und es säurehaltiger machte. Mehr zu essen führte auch dazu, dass die Korallen mehr Kohlendioxid ins Wasser ausstoßen. Das Ergebnis war ein starker Anstieg der Säure auf einen höheren Wert, als für die Zukunft aufgrund menschlicher Emissionen vorhergesagt wurde - dennoch florierte die Koralle weiter.
Diese Beobachtungen stehen im krassen Gegensatz zu der vorherrschenden Ansicht, dass ein Anstieg des Säuregehalts schädlich für Korallen ist, die zum Tode führen, wenn sie zu weit gehen. Aber die Werte, die von den Forschern mit dieser neuen Anstrengung gesehen wurden, deuten darauf hin, dass dies überhaupt nicht der Fall ist, und verwirren daher die Theorien über die Auswirkungen höherer Kohlendioxidwerte und wärmerer Temperaturen auf die Ozeane. Ein anderes Team aus Westaustralien bemerkte, dass die Ergebnisse, die von diesem neuen Team gefunden wurden, mit denen einer kleinen Studie übereinstimmten, die sie durchführten, wo sie Kisten um einige Korallen herum stellten und mit Kohlendioxid verrohrt wurden, ohne nachteilige Auswirkungen.
K. L. Yeakel et al. Veränderungen in der Biogeochemie des Korallenriffs und die daraus resultierende Versauerung im Zusammenhang mit der Offshore-Produktivität, Proceedings of the National Academy of Sciences (2015).
Bedeutung
Die Ozeanversauerung wird angenommen, um negative Auswirkungen auf die Ökosysteme der Korallenriffe zu haben, aber um zukünftige potenzielle Auswirkungen zu verstehen, ist es notwendig, die natürliche Variabilität und Kontrolle der Biogeochemie der Korallenriffe zu verstehen. Hier stellen wir eine 5-Jahres-Studie der Bermuda-Korallenriffplattform vor, die zeigt, wie schnelle interannuelle Versauerungsereignisse auf der lokalen Riffskala durch Verschiebungen in den biogeochemischen Prozessen des Riffs in Richtung einer Erhöhung der Netzkalkablagerung und der Netzatmung angetrieben werden. Diese biogeochemischen Veränderungen sind möglicherweise mit der Offshore-Produktivität verbunden, die letztendlich durch groß angelegte klimatologische und ozeanographische Prozesse gesteuert werden kann.
Abstract
Die ozeanische Aufnahme von anthropogenem Kohlendioxid (CO2) hat seit vorindustrieller Zeit das offen-ozeanische Oberflächenwasser um 0,1 pH-Einheiten versauert. Trotz eindeutiger Beweise für die Ozeanversauerung (OA) durch Messungen im offenen Meer in den letzten Jahrzehnten ist sie in Nearshore- und Korallenriffumgebungen noch nicht dokumentiert. Das Fehlen von Langzeitmessungen aus diesen Umgebungen schränkt unser Verständnis der natürlichen Variabilität und Kontrolle der CO2-Karbonat-Chemie und der Biogeochemie des Meerwassers ein, was für genaue Vorhersagen über die Auswirkungen zukünftiger OA auf Korallenriffe unerlässlich ist. Hier, in einer 5-jährigen Studie des Bermuda-Korallenriffs, zeigen wir Beweise dafür, dass Variationen in biogeochemischen Prozessen des Riffs zwischenjährliche Veränderungen des pH-Wertes des Meerwassers bewirken und Ωaragonite, die teilweise durch Offshore-Prozesse kontrolliert werden. In den Sommern 2010 und 2011 wurden schnelle Versauerungsereignisse beobachtet, die durch Verschiebungen in Richtung zunehmender Netzkalkablagerung und Netzheterotrophie getrieben wurden, wobei Häufigkeit und Ausmaß solcher Ereignisse einer erhöhten Offshore-Produktivität entsprachen. Diese Ereignisse fielen auch mit einem negativen North Atlantic Oscillation (NAO) Index zusammen, der historisch gesehen mit einer umfangreichen Offshore-Mischung und einer höheren Primärproduktivität am Standort Bermuda Atlantic Time-series Study (BATS) verbunden war. Unsere Ergebnisse zeigen, dass Korallenriffe natürlichen interannuellen Ereignissen der schnellen Versauerung durch Verschiebungen in den biogeochemischen Prozessen des Riffs ausgesetzt sind, die mit der Offshore-Produktivität verbunden sein können und letztendlich von größeren klimatischen und ozeanographischen Prozessen kontrolliert werden.
Die Ozeanversauerung (OA), die sich aus dem Anstieg des atmosphärischen CO2 (1⇓-3) und dem damit verbundenen Rückgang des pH-Wertes und des Sättigungszustands des Oberflächenmeerwassers in Bezug auf CaCO3-Mineralien wie Aragonit (Ωaragonite =[Ca2+][CO32-]/Ksp*, wobei Ksp* das Ionenlöslichkeitsprodukt ist) ergibt, hat Bedenken hinsichtlich der möglichen Folgen für marine Kalziumer und Ökosysteme aufgeworfen (4, 5). Reduzierungen auf Ωaragonite haben sich negativ auf die organismische CaCO3-Produktion ausgewirkt (6) und beschleunigen gleichzeitig die Bioerosion und die CaCO3-Auflösung (7, 8). Daher wurde angenommen, dass sich die Korallenriffe von einem Zustand der Netzkalkablagerung in eine Netzauflösung verschieben könnten, wobei einige Modellschätzungen einen Übergang für weltweite Riffe bei atmosphärischen CO2-Werten von 560 ppm vorhersagen (5, 7).
Trotz wachsender Besorgnis über die Anfälligkeit von Korallenriffen hat ein Mangel an Langzeitmessungen eine direkte Beobachtung der anthropogenen OA aufgrund des zunehmenden CO2-Ausstoßes in der Atmosphäre in diesen Umgebungen verhindert. Darüber hinaus haben kurzfristige Beobachtungen eine große Variabilität und Modifikation der CO2-Carbonat-Chemie des Riffwassers auf tages- und saisonaler Ebene als Ergebnis biogeochemischer Prozesse wie Photosynthese, Atmung, Verkalkung und CaCO3-Auflösung ergeben (9, 10). Es wurde angenommen, dass diese natürlichen Prozesse, quantifiziert als Netto-Ökosystemproduktion (NEP = Brutto-Primärproduktion - autotrophe und heterotrophe Atmung) und Netto-Ökosystemkalkung (NEC = Brutto-Kalkung - Brutto-CaCO3-Auflösung), die lokale Meerwasserchemie so modulieren könnten, dass sich die Versauerungsrate auf Korallenriffen deutlich vom offenen Ozean unterscheidet (11). Folglich könnte das anthropogene OA-Signal durch biogeochemische Prozesse im Riff entweder gemildert oder verschärft werden (10⇓-12). Jahrhunderte lange Aufzeichnungen über den pH-Wert des Riffs, die aus 11B Korallenkernen gewonnen wurden, zeigen eine große Variabilität des pH-Wertes über dekadische Zeitskalen (13), was möglicherweise auf die dynamische Natur biogeochemischer Prozesse im Riff hinweist. Diese Aufzeichnungen des pH-Wertes allein (die jedoch eher abgeleitet als direkt gemessen werden) können jedoch nicht explizit die Treiber hinter den beobachteten Schwankungen des pH-Wertes des Meerwassers aufdecken. Langzeitmessungen der Biogeochemie von Riffen (und Offshore) sind daher notwendig, um die natürlichen Variationen und Kontrollen (ob biologisch, physikalisch ozeanographisch oder klimatisch) der CO2-Carbonat-Chemie am Riff zu verstehen und wie sich diese unter zukünftigen OA- und Klimaszenarien ändern werden.
Wir haben die CO2-Carbonat-Chemie des Meerwassers auf der Bermuda-Korallenriffplattform monatlich zwischen Juni 2007 und Mai 2012 gemessen und charakterisiert, um die zeitliche und räumliche Variabilität von Meerwasser pCO2, pH und Ωaragonite zu untersuchen. Diese Parameter wurden basierend auf Oberflächenmeerwassermessungen von Temperatur, Salzgehalt, gelöstem anorganischem Gesamtkohlenstoff (DIC =[CO2]+[HCO3-]+[CO32-]) und Gesamtalkalität[TA = Überschuss an Basen über Säuren im Vergleich zu einem Referenzzustand (14)] an vier diskreten Stellen auf einem die Korallenriffplattform durchquerenden Transekt berechnet (Abb. S1). Angesichts der großen Variabilität der CO2-Systemparameter des Riffs über saisonale Zeiträume ist ein 5-jähriger Datensatz zu kurz, um den säkularen Trend der anthropogenen OA zu erkennen. Zeitgleiche monatliche Messungen der nahegelegenen Station Bermuda Atlantic Time-series Study (BATS), die sich ∼80 km südöstlich von Bermuda im offenen Ozean des subtropischen Gyros des Nordatlantiks befindet (Abb. S1), ermöglichen es uns jedoch, Variationen in der Offshore-Wasserchemie von Veränderungen am Korallenriff aufgrund lokaler biogeochemischer Prozesse zu entschlüsseln.
Fig. S1.
Bathymetrie der Korallenriffplattform der Bermudas mit Zeitreihenstudien. Riffzeitreihen werden durch weiße Felder gekennzeichnet (Tynes Bay, TB; Dock Yard, DY; Mid Platform, MP; North Channel, NC). An jeder Station wurden von Juni 2007 bis Mai 2012 monatlich Wasserproben zur Messung von DIC, TA, Temperatur und Salzgehalt entnommen. Der weiße Pfeil zeigt die allgemeine Richtung des BATS-Studienplatzes ∼80 km vor der Küste (31°50′ N, 64°10′W). Kredit: Mandy Shailer, Department of Conservation Services, Regierung der Bermudas.
Relative Veränderungen in DIC und TA spiegeln die biogeochemische Aufteilung des Kohlenstoffs zwischen den anorganischen und organischen Kohlenstoffkreisläufen am Riff (15, 16) und das Gleichgewicht von NEP, NEC und CO2-Gasaustausch in der See wider. NEC ändert DIC und TA in einem Verhältnis von 1:2, wobei die Netzkalkabscheidung (NEC > 0) dazu dient, DIC und TA abzusenken, was zu einer Senkung des pH-Wertes von Meerwasser und Ωaragonite führt. NEP verändert hauptsächlich DIC, wobei die Nettoproduktion von organischem Kohlenstoff (NEP > 0) die DIC-Konzentrationen reduziert, wenn CO2 verwendet wird, und den pH-Wert erhöht und Ωaragonite. Der CO2-Gasaustausch zwischen Luft und Meer wirkt sich nur auf die DIC aus, übt aber in der Regel einen geringen Einfluss auf die DIC in Bezug auf NEC und NEP in Riffumgebungen mit Verweilzeiten von wenigen Tagen aus, wie beispielsweise auf den Bermudas (17, 18). Folglich kann der Vergleich der salinitätsnormalisierten Veränderungen in Meerwasser DIC und TA (nDIC und nTA) zwischen dem Riff und der Küste verwendet werden, um den relativen Beitrag und die Variabilität in Riff NEC und NEP über Zeit und Raum zu bewerten. Abweichungen zwischen Riff und BATS nTA-Konzentrationen zeigen das relative Ausmaß des Riff-NEC, während Abweichungen in nDIC, korrigiert um die Einflüsse von NEC und CO2-Gasaustausch zwischen Luft und Meer, das relative Ausmaß des Riff-NEP anzeigen. In der vorliegenden Studie haben wir diese Beziehungen genutzt, um den Einfluss biogeochemischer Prozesse auf die Chemie des Riff-Meerwasserkarbonats und die relative Zuordnung von NEC und NEP zu Veränderungen des Riff-pH und Ωaragonite zu bewerten (siehe Materialien und Methoden für weitere Details).
Resultate und Diskusion
Langfristiges OA-Signal Offshore.
Beobachtungen seit 1983 bei BATS und der Hydrostation S (eine weitere langfristige Zeitreihe bei Bermudas) zeigen einen Anstieg des Oberflächenmeerwassers nDIC um ∼1.20 ± 0.09 μmol kg-1⋅y-1 (2), angetrieben durch die ozeanische Aufnahme von anthropogenem CO2 (Abb. 1). Infolgedessen ist das Oberflächenmeerwasser vor der Küste der Bermudas weniger alkalisch geworden, wobei der temperatur- und salznormalisierte pH-Wert und Ωaragonite seit 1983 (2) um -0,05 bzw. -0,25 Einheiten gesunken sind. (Beachten Sie, dass in dieser Diskussion auf temperatur- und salznormalisierte Daten verwiesen wird.) Der zunehmende nDIC hat die meisten dieser Veränderungen bewirkt, während nTA relativ konstant geblieben ist. Überlagert von dem stetigen Anstieg des nDIC im Laufe der Zeit ist eine saisonale Schwingung zu erkennen, die große Schwankungen des pH-Wertes und Ωaragonite verursacht. Dies zeigt den Einfluss physikalischer, biologischer und klimatischer Prozesse auf die Kohlenstoffdynamik des Oberflächenmeeres, wobei der vorherrschende Treiber saisonale Phytoplanktonblüten sind, wie sie im durchschnittlichen Jahreszyklus beobachtet werden.
Zeitreihen von Meerwasser nTA, nDIC, pH und Ωaragonite, die von 1983 bis 2014 bei BATS und Hydrostation S beobachtet wurden. (A) Langzeitbeobachtungen von nTA (blau) und nDIC (orange) zeigen eine relativ stabile nTA im Zeitverlauf, wobei der nDIC aufgrund der Aufnahme von anthropogenem CO2 zunimmt. Zeitliche Veränderung des pH-Wertes (B) und Ωaragonite (C), getrieben durch zeitabhängige Veränderungen in nTA (blauer Bereich) und nDIC (oranger Bereich). Die dicke schwarze Linie stellt die Gesamtveränderung sowohl des pH-Wertes als auch von Ωaragonite dar, wobei sich diese im Vergleich zu den ersten Beobachtungen ändert.
Riff- und BATS-Beobachtungen zur Meerwasser-Karbonatchemie, 2007-2012. (A-D) Zeitreihen von Meerwasser nTA, nDIC, pH und Ωaragonite, die von 2007 bis 2012 bei BATS (blau) und über die Riffplattform (orange) beobachtet wurden. Riffdaten werden als individuelle Marker pro Standort mit durchschnittlich vier Standorten als Linie mit ±1SD schattiert dargestellt; (E-H) Klimatologie der gleichen Meerwasser-CO2-Parameter mit ±1SD schattiert. Schnelle Versauerungsereignisse zeigen sich in den kurzfristigen Beobachtungen am Riff (A-D), insbesondere im Sommer 2010. Die Klimatologie zeigt, dass die Sommerzeit im Riff nTA und nDIC sinkt, während der pH-Wert im Riff und Ωaragonite das ganze Jahr über relativ konstant bleiben. Der pH-Wert und Ωaragonite wurden auf Werte von 23,1 °C bzw. 36,6 g kg-1 sowohl für BATS- als auch für Riffdaten normiert.
Near-Shore biogeochemische Prozesse diktieren die Versauerung von Riffen.
Im 5-jährigen Untersuchungszeitraum zwischen 2007 und 2012 werden auf der Korallenriffplattform keine sichtbaren Trends bei den Parametern der Seewasser-CO2-Carbonat-Chemie beobachtet, die sich aus dem Anstieg des atmosphärischen CO2 ergeben (Abb. 2). Allerdings werden jedes Jahr große Verarmungen in nTA und nDIC im Vergleich zu BATS beobachtet, wobei das Ausmaß der Drawdown-Variablen auf der interannuellen Zeitskala berücksichtigt wird. Die Offshore-Variabilität der Seewasser-Karbonat-Chemie bei BATS ist repräsentativ für die Bedingungen im subtropischen Gyre des Nordatlantiks um Bermudas herum und bietet einen direkten Vergleich mit der Variabilität an Land am Bermuda-Riff. Trotz dieser Erschöpfung des Riffs nTA und nDIC bleiben der pH-Wert des Riffs und Ωaragonite vergleichsweise konstant, während vor der Küste ein vorhersagbares saisonales Muster der Versauerung während der gesamten 5-y-Studie anhält (Abb. 2).
Gelegentlich werden auf der Riffplattform schnelle Versauerungsereignisse beobachtet, insbesondere im August 2010, als der größte Unterschied im pH-Wert zwischen Riff und BATS (ΔpHREEF-BATS = -0,14, Abb. 3) beobachtet wurde. Die Klimatologie des pH-Wertes des Riffs und Ωaragonite zeigt, dass der Variabilitätsbereich größer ist und die Durchschnittswerte im Sommer im Vergleich zum Rest des Jahres etwas niedriger sind, was mit dem Rückgang und der maximalen Variabilität von nTA und nDIC zusammenfällt (Abb. 2 E-H).
Einfluss der NAO- und Offshore-Primärproduktivität auf die Dynamik der Riffkarbonatchemie. Zeitreihen von (A) NAO[monatlich in Grau und Winter (Dezember-März), Mittelwert in Schwarz]; (B) Windgeschwindigkeit (grün) und Mischschichttiefe (MLD) (schwarz); (C) Primärproduktion (PP); nTA (D) und nDIC (E) bei BATS (Felder) und über die Riffplattform (Kreise); (F) NEC (blau), NEP (rot) und CO2-Gasaustausch in der See (grün); und (G) die Beiträge von NEC, NEP und Gasaustausch in der See zu pH-Differenzen zwischen BATS und dem Riff (ΔpHREEF- BATS). Negative NAO-Winterereignisse verstärken die Sturmaktivität in subtropischen Gewässern um die Bermudas herum und vertiefen die MLD, die kälteres, relativ nährstoffreiches Wasser an die Oberfläche bringt und zu mehr PP führt. In den Jahren 2010 und 2011 fiel die MLD im tiefen Winter mit verstärkten Frühlingsblüten und einer verbesserten NEC im Spätsommer/Herbst zusammen, was zu einem stärkeren Absinken von nTA im gesamten Riff führte und somit die Versauerung vorantrieb. Im Gegensatz dazu führte ein positiver Winter (Dezember-März) NAO-Zustand und damit ein schwächeres PP-Signal im Jahr 2008 zu einem gedämpften Riff-NEC-Signal im folgenden Sommer.
Der beobachtete Rückgang im Riff nTA wird hauptsächlich durch das Riff NEC verursacht, wobei die Modifikationen im nDIC sowohl durch das Riff NEC als auch durch das NEP getrieben werden (die Einflüsse des Luft-/Seegasaustauschs von CO2 sind minimal, Abb. 3). Folglich treiben saisonale Veränderungen in NEC und NEP letztendlich die beobachtete pH- und Ωaragonite-Variabilität am Riff voran, wobei die zunehmende NEC die Versauerung verursacht und die zunehmende NEP den gegenteiligen Effekt hat. Erhöhungen der Sommerzeit in NEC führen zu einer wiederkehrenden Senkung des pH-Wertes des Meerwassers, wobei die Stärke von NEC dem Ausmaß der pH-Senkung entspricht (Abb. 3). Im Gegensatz dazu schwankt die NEP ohne erkennbaren saisonalen Zyklus. Wenn ein Anstieg der NEC mit einem Rückgang der NEP korrespondiert, wie im Sommer 2010, beobachten wir die stärkste Senkung des pH-Wertes von Riff-Meerwasser und Ωaragonite (Abb. 3).
Die Biogeochemie des Riffs reagiert auf den Offshore-Zwang.
Unsere Ergebnisse zeigen, dass saisonale und interannuelle Veränderungen in NEC und NEP für die beobachteten Veränderungen des temperatur- und salznormalisierten pH-Wertes und Ωaragonite verantwortlich sind, aber was ist der Treiber dieser Veränderungen? Ausgeprägte Versauerungsereignisse, wie z.B. im Sommer 2010, traten in Perioden mit anomal hohem NEC und niedrigem NEP auf, was auf Verschiebungen in biogeochemischen Prozessen im Riff hin zu erhöhter Verkalkung und Heterotrophie hinweist. Im Allgemeinen wird die Verkalkung als ein autotrophisch verstärkter Prozess betrachtet, bei dem die photosynthetische Absenkung der CO2-Konzentrationen im Meerwasser Ωaragonite erhöht und günstige Bedingungen für die organismische CaCO3-Ablagerung schafft (19). Im Gegensatz dazu setzt die Atmung CO2 frei, reduziert Ωaragonite und soll daher die Verkalkung verringern. So steht unser Ergebnis einer verstärkten Verkalkung in Zeiten erhöhter Heterotrophie im Widerspruch zu einigen Modellierungs- und Feldstudien (10, 19). Zahlreiche Experimente haben jedoch gezeigt, dass Korallen, die dominierenden Riffkalker, gut gefüttert werden (z.B. durch Zooplankton) oder eine nährstoffreiche Ernährung haben, sowohl Gewebe- als auch Skelettwachstumsraten aufweisen werden (20⇓⇓⇓-24). Einige Ergebnisse haben sogar gezeigt, dass Korallen in der Lage sein könnten, reduzierten Verkalkungsraten aufgrund von OA durch eine Erhöhung der Fütterungsraten entgegenzuwirken (23). Darüber hinaus wurde festgestellt, dass eine erhöhte Heterotrophie und ein erhöhtes Gewebewachstum bei Korallen dazu führen, dass die Gesamtatemraten steigen (21). Daher spekulieren wir, dass externe Nährstoffimpulse an das Riff sowohl die anomal hohe Sommerkalkablagerung als auch Verschiebungen zur zunehmenden Heterotrophie, die für die beobachteten Versauerungsereignisse verantwortlich ist, hätten ermöglichen können.
Messungen bei BATS deuten darauf hin, dass die Phytoplanktonblüten während unseres Untersuchungszeitraums verbessert wurden, insbesondere im Winter und Frühjahr 2010 und in geringerem Maße auch 2011 (Abb. 3). Während die Sargasso-See um die Bermudas herum typischerweise oligotroph ist, wurden bereits erhöhte Frühlings-Phytoplanktonblüten dokumentiert und mit der Nordatlantischen Oszillation (NAO) (25, 26) in Verbindung gebracht, einem Klimamodus, der Druckunterschiede zwischen dem atmosphärischen subtropischen Hochdrucksystem nahe den Azoren und dem subpolaren Niederdrucksystem nahe Island beschreibt (27). Ein negativer Winter NAO-Zustand führt zu einer südlichen Verschiebung des Golfstroms und der Wintersturmspur, was aufgrund verbesserter Winde zu tieferen Mischschichten im Sargassomeer führt (25, 27). Mit tieferen gemischten Schichten kommen niedrigere Meeresoberflächentemperaturen und eine stärkere Nährstoffaufnahme, was zu Produktivitätsblüten und einem erhöhten Mesozooplankton-Überschuss führt (25, 26, 28⇓-30).
Seit Beginn der Messungen bei BATS war der Winter NAO-Zustand in erster Linie positiv, wobei sporadisch neutrale bis leicht negative Ereignisse zu dokumentierten Produktivitätssteigerungen auf See führten (25, 26). Das Ereignis im Winter 2010 (und in geringerem Maße auch 2011) war jedoch anomal negativ (Abb. 3) und fiel mit einer ungewöhnlich großen Frühjahrsblüte zusammen. Zu Beginn des Ereignisses 2010 fiel eine stark negative Verschiebung der Riff-NEP (d.h. der Nettoatmung) mit einer tiefen Mischschichttiefe (MLD) und einem daraus resultierenden Auftrieb von Nährstoffen zusammen[was durch eine anomal niedrige Meeresoberflächentemperatur (SST) und hohe nDIC-Wasser bei BATS angezeigt wird, Abb. S2-S4]. (25). Reef nTA-Werte, die über denen bei BATS lagen, deuten darauf hin, dass die Verschiebung in der NEP mit der plattformweiten Netto-CaCO3-Auflösung zusammenfiel (Abb. 3). Während die erhöhte Offshore-Primärproduktivität im Winter bis in die Frühjahrssaison 2010 und 2011, im folgenden Spätsommer und Herbst, auftrat, erreichten die relativen Messungen des Riff-NEC, wie sie durch Alkalinitätsanomalien abgeleitet wurden, ihre höchsten Werte über den 5-jährigen Messzeitraum, was eine verstärkte Verkalkung der Gemeinschaft bedeutet (max. NEC 2007): 39,8 μmol kg-1; 2008: 30,9 μmol kg-1; 2009: 42.2 μmol kg-1; 2010: 50,5 μmol kg-1; 2011: 50,0 μmol kg-1, Abb. 3), und fiel mit negativen NEP-Werten zusammen. Bemerkenswert ist, dass in den folgenden Wintern 2011 und 2012 auch die NEC-Raten im Vergleich zu den Vorjahren höher waren (Abb. 3). Die Kreuzkorrelationsanalyse ergab die potenzielle Zeitabhängigkeit zwischen Offshore- und Riffprozessen, wobei die maximale Korrelation zwischen Offshore-Primärproduktivität und MLD beobachtet wurde, wenn die Produktivität um 0 bis 1 mo hinter MLD zurückblieb (r > 0,8, Abb. 4). Im Gegensatz dazu verzögerten biogeochemische Prozesse im Riff die Offshore-Produktivität um mehrere Monate. Reef NEC zeigte die stärkste Korrelation mit der Offshore-Produktivität bei einer Verzögerung von 4 mo (r = 0,77), während negative NEP (d.h. Heterotrophie) bei einer Verzögerung von 6 mo (r = -0,45, Abb. 4) am stärksten korreliert waren. Diese Korrelationen waren bei einem Konfidenzniveau von 99,5% statistisch signifikant.
Mehr aus dieser Studie finden Sie hier

{kind=link}
{kind=link}
{kind=link}
{kind=link}