- Tais W. Dahl,
- Magnus A. R. Harding,
- Julia Brugger,
- Georg Feulner,
- Kion Norrman,
- Barry H. Lomax &
- Christopher K. Junium
Nature Communications volume 13, Article number: 7616 (2022) Cite this article
Abstract
Die Entstehung der Wälder auf der Erde (vor ca. 385 Millionen Jahren)1 wurde mit einem Rückgang des atmosphärischen CO2-Gehalts um eine Größenordnung und einer globalen Klimaabkühlung durch Veränderung der kontinentalen Verwitterungsprozesse in Verbindung gebracht, aber die Beobachtungen zum atmosphärischen CO2-Gehalt vor der Entstehung der Wälder sind mit großen, oft ungebundenen Unsicherheiten behaftet. Hier kalibrieren wir ein mechanistisches Modell für den Gasaustausch in modernen Lycophyten und bestimmen den atmosphärischen CO2-Gehalt von 410-380 Ma anhand verwandter fossiler Pflanzen mit einer Unsicherheit von etwa ±100 ppm (1 sd). Wir stellen fest, dass die Atmosphäre ~525-715 ppm CO2 enthielt, bevor die Kontinente aufgeforstet wurden, und dass die Erde nach einem Paläoklimamodell teilweise vergletschert war. Ein prozessgesteuertes biogeochemisches Modell (COPSE) zeigt, dass das Auftreten von Bäumen mit tiefen Wurzeln den CO2-Abbau in der Atmosphäre nicht dramatisch erhöht hat. Vielmehr könnten flachwurzelnde vaskuläre Ökosysteme lange vor dem Aufkommen der Wälder gleichzeitig eine abrupte Sauerstoffanreicherung der Atmosphäre und eine Abkühlung des Klimas verursacht haben, obwohl die früheren CO2-Werte noch unbekannt sind.
Einleitung
CO2 in der Atmosphäre ist ein Spurengas, das das Klima der Erde im Laufe der Erdgeschichte beeinflusst hat2,3. Seine Schwankungen in der Vergangenheit geben uns Aufschluss über die natürlichen langfristigen Quellen und Senken. In Abwesenheit der anthropogenen Verbrennung fossiler Brennstoffe ist die wichtigste atmosphärische CO2-Quelle die vulkanische Ausgasung, und diese Quelle wird hauptsächlich durch den Abbau ausgeglichen, der erfolgt, wenn CO2-haltige Flüssigkeiten chemisch reagieren und Silikatgestein verwittern, gefolgt von der Ablagerung von Karbonat in den Ozeanen4. Die Auflösung von Silikatmineralien in der Verwitterungszone erfolgt durch Wechselwirkungen zwischen dem terrestrischen Ökosystem und geologischen Prozessen, die frisches Gestein an der Oberfläche für die Reaktion verfügbar machen. Die Rolle der Biologie und die CO2-Empfindlichkeit der Rückkopplungen, die den globalen CO2-Abbau steuern, sind jedoch umstritten5,6,7,8,9. Es wird vermutet, dass die verstärkte kontinentale Verwitterung einen Rückgang des atmosphärischen CO2-Drucks (pCO2) von einem Niveau, das etwa zehnmal höher war als die heutige Konzentration3,10,11, auf ein annähernd modernes Niveau verursacht hat, das mit dem devonisch-karbonischen Übergang von Treibhaus- zu Eishausbedingungen als Reaktion auf die Aufforstung der Kontinente verbunden ist. Jüngste geochemische Nachweise und Erdsystemmodelle12 deuten jedoch darauf hin, dass die atmosphärische Sauerstoffanreicherung lange vor der Entwicklung der Bäume auf den Kontinenten stattfand (~393-383 Ma1,3). Außerdem ist der zeitliche Zusammenhang zwischen der Besiedlung durch Pflanzen und der permokarbonischen Vergletscherung umstritten8. Es gibt zwingende Beweise dafür, dass die Erde im Ordovizium-Silur ebenfalls in einen vergletscherten Zustand überging13. Der Zusammenhang zwischen Vergletscherung und atmosphärischem CO2 ist jedoch kompliziert, und Paläoklimamodelle zeigen, dass die Vergletscherung selbst bei einem 12-14-fachen des vorindustriellen atmosphärischen Niveaus (PIAL, 280 ppmv) andauern könnte14. Daher ist eine genaue Rekonstruktion des atmosphärischen pCO2-Wertes im Zusammenhang mit der Pflanzenentwicklung von entscheidender Bedeutung, um die Auswirkungen der terrestrischen Biota auf das Klima der Erde zu bewerten. Hier zeigen wir, dass der atmosphärische pCO2-Wert deutlich niedriger war als bisher angenommen, als Bäume und Wälder auf unserem Planeten auftauchten.
Nach kanonischer Auffassung lagen die atmosphärischen CO2-Konzentrationen im frühen Paläozoikum um eine Größenordnung über den vorindustriellen Werten3,10,15,16,17, obwohl neuere Studien von Werten ausgehen, die viel näher an den heutigen liegen18. Traditionell sind Paläo-CO2-Schätzungen aus Proxydaten mit großen und manchmal unbegrenzten Unsicherheiten verbunden19,20. Für das späte Ordovizium (~445 Ma) gibt es beispielsweise Belege für hohe pCO2-Werte von 17 ± 4 PIAL (1 sd, Standardabweichung) aus CO2, das in pedogenem Goethit aus der Neda-Formation in Wisconsin, USA, enthalten ist19. Dieses Paläosol zeigt eine Kopplung zwischen dem CO2-Gehalt im Goethit und seiner Isotopenzusammensetzung, die als Ausdruck einer variablen Vermischung von atmosphärischem CO2 und in das Goethit-Mineralgitter eingelagertem, vom Boden aufgenommenem Kohlenstoff interpretiert wird. Die obige Spanne spiegelt alle analytischen Fehler wider, die in die Berechnung eingeflossen sind (siehe Einzelheiten in den ergänzenden Informationen, SI), und wir weisen darauf hin, dass der Fehler noch größer sein könnte, wenn die CO2-Oberflächenadsorptionseigenschaften21 auf natürlichem Goethit von denen des im Labor gezüchteten phosphatierten Goethits22 abweichen. Dies wurde in modernen Böden noch nie nachgewiesen, und der Proxy wurde auch noch nie zu einem anderen Zeitpunkt der Erdgeschichte angewandt.
Außerdem wurde aus der Kohlenstoffisotopenzusammensetzung des pedogenen Karbonats ein systematischer Rückgang des atmosphärischen CO2-Gehalts im Devon von 5 auf 0,7 PIAL abgeleitet2,16,17. Obwohl der rückläufige Trend real sein könnte, wurden die absoluten atmosphärischen CO2-Werte, die aus pedogenen Karbonaten aus diesem Zeitintervall gemeldet wurden, weiter nach unten korrigiert23 und sind wahrscheinlich systematisch überschätzt, da die Produktivität in den Böden des frühen Paläozoikums im Vergleich zu den modernen Böden geringer ist17. Die Weitergabe der Unsicherheit, die mit angenommenen Modellparametern verbunden ist, die nicht unabhängig von den Gesteinsaufzeichnungen bestimmt werden können, wie z. B. der Anteil des vom Boden aufgenommenen CO2 im Boden und seine Isotopensignatur, zeigt, dass die auf diese Weise erhaltene absolute Paläo-pCO2-Schätzung mit einer Unsicherheit von einer Größenordnung behaftet ist (siehe ergänzende Abb. 15 und die ergänzenden Informationen für Details).
Darüber hinaus gibt es erste Hinweise auf hohe paläozoische CO2-Werte (~16 PIAL)20 , die sich aus der geringen Stomatendichte (mm-2) einiger fossiler Pflanzen (z. B. Aglaophyton, Sawdonia) ergeben. Diese anatomischen Merkmale wurden als Beweis dafür gedeutet, dass sich diese Pflanzen an die Minimierung des Wasserverlustes in einer Atmosphäre mit hohem CO2-Gehalt angepasst haben, aber es ist wichtig, sie mit Pflanzen zu vergleichen, die eine ähnliche Anatomie des Gasaustausches und eine ähnliche Verhaltenskontrolle über die Wasserverlustrate und die CO2-Aufnahme aufweisen. Dies ist bei dieser Gruppe rätselhafter früher Gefäßpflanzen, von denen es keine lebenden Nachkommen gibt, sehr problematisch. Im Vergleich dazu weisen Fossilien von Lycophyten, die mit diesen ausgestorbenen Pflanzengruppen zusammen vorkommen und die moderne Verwandte mit ähnlicher Physiologie haben, eine ähnliche Stomatendichte auf wie ihre modernen Nachkommen (ergänzende Tabelle 5), was darauf hindeutet, dass die von Aglaophyton- und Sawdonia-Exemplaren abgeleiteten hohen CO2-Vorhersagen falsch sein könnten und dass der atmosphärische CO2-Gehalt im frühen Paläozoikum viel näher am modernen Niveau lag24.
Kürzlich wurde festgestellt, dass der atmosphärische CO2-Gehalt im späten Ordovizium (~400-700 ppm) und im mittleren Devon (~700-1400 ppm) nur geringfügig erhöht war, und zwar auf der Grundlage von Kohlenstoffisotopendaten des marinen Phytoplanktons, die über ein diagenetisches Produkt des Chlorophylls, das Phytan, erfasst wurden18. Zwischen ~432 Ma und 390 Ma werden keine Phytan-CO2-Daten gemeldet (ergänzende Abbildungen 19-21). Der Phytan-Proxy zeigt die erwartete Reaktion bei erhöhten CO2-Konzentrationen und sagt sinkende CO2-Werte in Abhängigkeit von der Entfernung zu einem modernen CO2-Austrittsort voraus25 (wenn auch mit größeren Fehlern als bei paläozoischem Phytan). Bei Anwendung auf modernes Phytoplankton sagt der Phytan-Proxy auch heute noch eine große Bandbreite atmosphärischer CO2-Konzentrationen voraus (~300-1200 ppm)26. Die Genauigkeit dieses Proxys hängt von Variablen ab, die sich nicht ohne weiteres aus den geologischen Aufzeichnungen ablesen lassen, darunter das Verhältnis von atmosphärischem CO2 zu gelöstem CO2 des Meerwassers, in dem das Phytoplankton wuchs, die Art der Phytoplanktonarten, andere Phytanquellen, die Wachstumsrate des Phytoplanktons und damit die isotopische Reaktion des Phytoplanktons, von dem das Phytan abgeleitet wurde, auf die Konzentration des gelösten CO2 in der Umgebung26. In Anbetracht der vielen Unsicherheiten sind die mit dem Phytan-CO2-Proxy verbundenen Fehler wahrscheinlich groß.
Nach demselben Grundprinzip und mit der Möglichkeit, alle beteiligten Parameter einzuschränken, reagiert die Kohlenstoffisotopenfraktionierung in Pflanzengewebe von Landpflanzen (Δleaf) empfindlich auf die interne CO2-Konzentration in den substomatären Hohlräumen, die wiederum von der Wasserverfügbarkeit und dem umgebenden pCO2 in der Umwelt abhängt27. Kürzlich hat die Kombination von Isotopendaten und Stomata-Parametern zur Entwicklung eines mechanistischen Ansatzes für die Vorhersage des Paläo-pCO2 geführt, der auf dem Gasaustausch zwischen Blättern basiert27. Die Anwendung dieses Ansatzes auf die fossilen Aufzeichnungen zeigt, dass der atmosphärische CO2-Gehalt nach dem Devon die meiste Zeit
Wichtig ist auch, dass mit diesem mechanistischen Ersatzmodell sowohl die Genauigkeit als auch die Präzision der Vorhersage getestet werden konnte, so dass realistische Einschränkungen für den vorhergesagten Paläo-pCO2 bewertet werden können. So ermöglicht der Rahmen, der dem Proxy zugrunde liegt, eine Neubewertung der CO2-Vorhersagen von Langzeit-Kohlenstoffkreislaufmodellen, deren Vorhersagen mit großen Unsicherheiten behaftet sind11,12,15 , sowie der Vorhersagen von Paläo-pCO2-Proxies der ersten Generation, die weitgehend unkontrollierte pCO2-Schätzungen lieferten.
Das mechanistische Modell von Franks et al.27 verwendet zur Vorhersage des paläo-pCO2 sowohl Kohlenstoffisotopendaten als auch stomatäre Merkmale (Dichte und Größe), die aus den Fossilien gewonnen wurden. Beide Merkmale sind ein Ausdruck dafür, wie die Pflanze zu Lebzeiten an ihre lokale Umgebung angepasst war. Folglich gibt es eine enge Beziehung zwischen diesen verschiedenen "Aufzeichnungsgeräten" für die Wechselwirkungen zwischen Pflanzen und Klima. So wurde beispielsweise nachgewiesen, dass die Kohlenstoff-Isotopensignatur des Blattgewebes stark von der Wasserverfügbarkeit beeinflusst wird28 und dass die Nützlichkeit von mechanistischen Proxies, die sich ausschließlich auf Isotopendaten stützen29,30,31 , in Frage gestellt werden muss32,33,34,35,36. Aufgrund der wechselseitigen Abhängigkeit von Isotopen- und Stomata-Parametern wird empfohlen, diese Daten nach Möglichkeit aus derselben Probe oder aus zeitgleichen Sedimentablagerungen zu gewinnen. Mit diesem Ansatz sollte sich die Genauigkeit der Vorhersage verbessern, da er gleichzeitig die variable CO2-Assimilationsrate und den Wasserleitwert als Reaktion auf die sich ändernde Wasserverfügbarkeit berücksichtigt27. Hier kalibrieren wir ein Blattgasaustauschmodell für Lycophyten und schätzen den atmosphärischen CO2-Gehalt mit Unsicherheiten, indem wir es auf einige der ältesten Vertreter aus dem Fossilarchiv anwenden.
Ergebnisse und Diskussion
Kalibrierung eines robusten Paläo-CO2-Barometers für Lycophyten
Um die frühen atmosphärischen CO2-Aufzeichnungen zu verankern, haben wir ein mechanistisches Modell des Blattgasaustauschs27 an lebenden Vertretern der ältesten Gefäßpflanzen-Taxa (Lycopodiaceae) kalibriert, das durch ihre Stomatendichte, Spaltöffnungsgröße und Kohlenstoffisotopenzusammensetzung eingeschränkt ist, und es auf fossile Überreste von Lycophyten aus dem Unter- und Mitteldevon mit der gleichen Stomatanatomie37 angewendet. Wenn man versucht, die fossilen Pflanzenfunde zu nutzen, um die CO2-Konzentration im unteren Paläozoikum zu dem Zeitpunkt abzuschätzen, als die Gefäßpflanzen ihre erste Ausbreitung erlebten, ist es eine Herausforderung, gepaarte Sätze von Stomata- und Isotopendaten zu verwenden. Diese Situation ergibt sich aus der Art der Erhaltung der Fossilien, der Notwendigkeit einer zerstörenden Analyse für Kohlenstoffisotope und dem wissenschaftlichen Wert der kuratierten Exemplare. Daher haben wir das mechanistische Modell27 weiterentwickelt, um eine Solver-Routine einzubauen, die es ermöglicht, für jeden der beobachtbaren Parameter, die für die Erstellung von Paläo-pCO2-Schätzungen erforderlich sind, einen breiten Bereich zu propagieren. Dieser Ansatz wurde entwickelt, um alle Variationen zwischen lokalen Umgebungen und/oder eine zeitliche Diskrepanz zwischen Datenquellen in die Modellvorhersagen einfließen zu lassen. Um zu überprüfen, ob unser Solver-Ansatz vernünftige Schätzungen des Paläo-pCO2 liefert, haben wir die Technik auf unseren vorhandenen Lycophyten-Datensatz angewandt.
Zunächst kalibrierten wir das Gasaustauschmodell27 anhand von zwei Lycophytenarten (Huperzia phlegmaria und H. squarrosa), die physiologisch vermutlich den Lycophyten aus dem Devon ähneln und bei bekannten CO2-Werten in der Umgebung über einen Zeitraum von etwa 8 Jahren bei natürlichem Licht in einem Bodensubstrat und bei einer optimalen relativen Luftfeuchtigkeit von etwa 80 % in einem Gewächshaus des Botanischen Gartens in Kopenhagen gewachsen sind. Messungen der Blatt-Kohlenstoff-Isotopenfraktionierung (Δleaf), der stomatären Dichte (SD) und der stomatären Porenlänge (p) (ergänzende Tabelle 2; ergänzende Daten 1) deuten darauf hin, dass ihre CO2-Assimilationsrate bei Referenz-CO2 (A0) nicht von der anderer moderner Lycophyten zu unterscheiden war (A0 = 3,7 ± 1,6 μmol m-2 s-1, 6 Arten). Unter Verwendung dieses durchschnittlichen A0-Wertes als repräsentativ für Lycophyten und unter Beibehaltung anderer Parameter, wie z. B. der operationellen stomatären Leitfähigkeitseffizienz (ζ) mit geringem Einfluss auf die pCO2-Schätzungen (siehe ergänzenden Text für Details), konnten wir die im Gewächshaus gemessenen CO2-Werte von 448 ± 51 ppm (tägliche Schwankung, 1 sd) mit einer CO2-Vorhersage von 407+24-22407-22+24 ppm bzw. 545+69-56545-56+69 ppm (Median ± Quartile) für die beiden verschiedenen Lycophytenarten reproduzieren.
Im Anschluss an dieses erste Screening untersuchten wir das mechanistische Modell zur Ableitung von CO2-Schätzungen weiter, indem wir einen Datensatz entwickelten, der auf nicht gepaarten Daten basierte, d. h. bei dem die Spaltöffnungen und Isotopendaten von H. phlegmaria und H. squarrosa aus unterschiedlichen Wachstumsbedingungen stammen, um fossile Proben zu imitieren, die aus geografisch/umweltmäßig unterschiedlichen Sedimentablagerungen ähnlichen Alters stammen. Basierend auf kultivierten Lycophyten, die unter trockenen und feuchten Bedingungen gewachsen sind, stellen wir fest, dass ungepaarte Daten mit bis zu ~100 ppm zusätzlicher Unsicherheit behaftet sein könnten, wenn Stomatendaten und Isotopendaten von Pflanzen stammen, die unter sehr unterschiedlichen Bedingungen gelebt haben; d. h. Stomatendaten von langsam wachsenden oder sogar halb-epiphytischen Pflanzen unter extrem trockenen Bedingungen mit Isotopendaten von Pflanzen, die unter optimalen, feuchten Bedingungen gewachsen sind oder umgekehrt (ergänzende Daten 1). Bei der Untersuchung dieses Datenrahmens zeigen die revidierten Vorhersagen von pCO2 eine gute Übereinstimmung sowohl mit dem tatsächlichen Wachstums-CO2 als auch mit den CO2-Werten, die mit der Standardmodellparametrisierung vorhergesagt wurden (Abb. 1-2; ergänzende Daten 1). Somit sollte die gleichzeitige Analyse von fossilem Lycophyten-ΔBlatt, SD und p sowohl genaue als auch präzise Vorhersagen von Paläo-pCO2 liefern, unabhängig davon, ob die Daten aus gepaarten Proben (Stomata- und Isotopendaten von der gleichen Lokalität) oder aus verschiedenen Lokalitäten stammen. Durch die Anwendung des mechanistischen Modells27 auf Lycophyten-Daten sind wir nun zum ersten Mal in der Lage, den atmosphärischen pCO2-Wert vor der Entstehung der Wälder genau und präzise zu bestimmen.
Abb. 1: Der atmosphärische pCO2-Gehalt wird aus der Isotopenfraktionierung des Blattkohlenstoffs (Δleaf), der Spaltöffnungsdichte (SD) und der Porenlänge der Spaltöffnungen (p) abgeleitet.
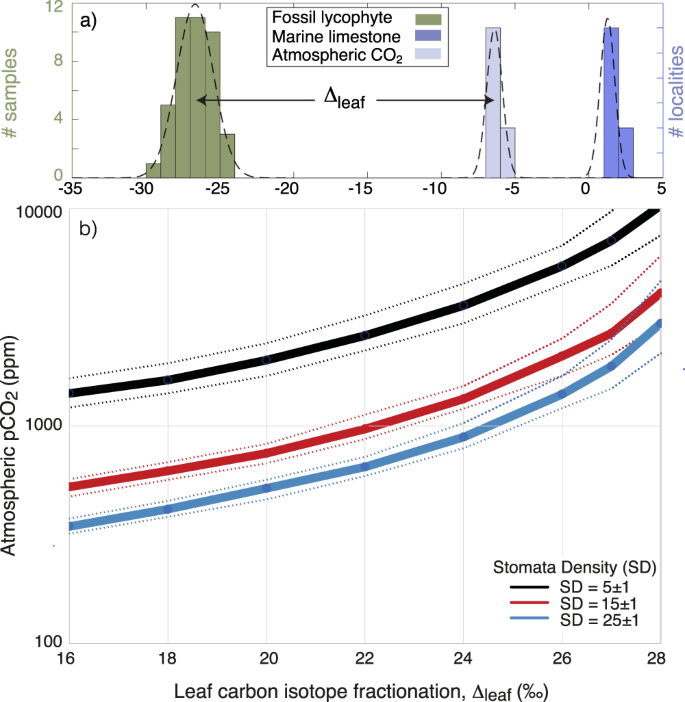
a Das ΔBlatt wird aus der Kohlenstoffisotopenzusammensetzung (δ13C) der fossilen Flora (grün), der marinen Karbonate (blau) und des berechneten atmosphärischen CO2 (hellblau) bestimmt (ergänzende Tabelle 4). b Pflanzen, die unter höheren CO2-Werten in der Umgebung gewachsen sind, weisen ein höheres ΔBlatt und/oder eine geringere Spaltöffnungsdichte auf. Die Kurven stellen ein Gasaustauschmodell dar, das für moderne Lycophyten mit einer Referenz-CO2-Assimilationsrate bei modernen atmosphärischen Werten (A0) von 3,5 µmol m-2 s-1, einer operativen stomatären Leitfähigkeitseffizienz (Verhältnis von operativer zu maximaler stomatärer Leitfähigkeit) ζ = 0,2, einer Stomata-Porenform β = 0,6 und p = 18 ± 2 µm kalibriert wurde, die mit alten und modernen Lycophyten vergleichbar sind (für Details siehe Anhang). Hier wird ein fossiler Lycophyt aus dem Devon als Beispiel verwendet. Lycophyten aus dem Devon weisen typischerweise Δ-Blatt-Werte nahe 20 und Spaltöffnungsdichten zwischen 15 und 25 mm-1 auf (Supplementary Data 1).
Abb. 2: Fehlerfortpflanzung des mechanistischen pCO2-Proxys, angewandt auf Givetian Drepanophycus sp. aus der Hamilton Group, NY, USA.
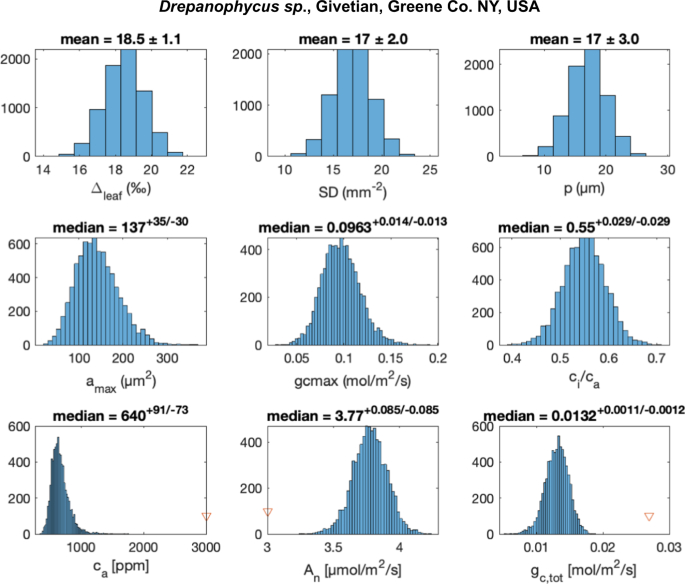
Wahrscheinlichkeitsverteilungen für Modelleingabeparameter (Δleaf, SD, p; obere Reihe) werden abgetastet (N = 10.000), um Modellparameter (mittlere Reihe: Spaltfläche, amax, maximaler Leitwert bei vollem Tageslicht, gc,max, und Verhältnis der CO2-Konzentration in den substomatären Hohlräumen zum atmosphärischen CO2, ci/ca😉 und posteriore Wahrscheinlichkeitsverteilungen für die Modellausgabeparameter (untere Reihe: atmosphärisches CO2, ca, CO2-Assimilationsrate An und Gesamtleitwert, gc,tot) zu berechnen. Die numerischen Lösungen wurden mit der Matlab-Funktion fsolve ermittelt (siehe Einzelheiten in den zusätzlichen Informationen). Die Ausgangsparameter der ersten Schätzung sind durch rote Dreiecke gekennzeichnet, und die Ergebnisse hängen nicht von der ersten Schätzung ab. Die Medianwerte und Fehler, die durch das 1. und 3. Quartil dargestellt werden, sind über jedem Feld für die berechneten Eigenschaften angegeben.
Der atmosphärische CO2-Gehalt in der Zeit von 410,8 bis 382,7 Ma (Pragian bis Givetian) wurde aus 66 fossilen Lycophyten rekonstruiert, die drei verschiedene Gattungen (Asteroxylon, Baragwanathia und Drepanophycus) aus 13 geologischen Ablagerungen an neun geographisch unterschiedlichen Orten repräsentieren. Daraus leiten wir vier pCO2-Schätzungen aus gepaarten Daten und sechs Vorhersagen aus ungepaarten Daten ab (ergänzende Daten 1 für Standort- und Datendetails). Wir stellen jedoch fest, dass die Spaltöffnungsdichten an allen Standorten bemerkenswert ähnlich waren (±1,5 mm-2, 1 sd) und auch die Δ-Blätter eine gewisse Ähnlichkeit aufweisen (18,3 ± 1,1, 1 sd). Bei Verwendung gepaarter Daten liegen die pCO2-Vorhersagen zwischen 525+139-101525-101+139 ppm und 695+99-73695-73+99 ppm (Median ± Quartile der Wahrscheinlichkeitsverteilung). Ungepaarte Vorhersagen ergeben einen Bereich von pCO2-Schätzungen, der zwischen 532+77-76532-76+77 ppm und 715+140-102715-102+140 ppm liegt. Unsere Daten liefern konsistente CO2-Schätzungen, unabhängig von der Methode, die für die Zusammenstellung der Rohdaten zur Unterstützung der Vorhersagen verwendet wurde (Abb. 3; Ergänzende Daten 1). Wir kommen daher zu dem Schluss, dass diese Daten belegen, dass der atmosphärische CO2-Gehalt während des ~30-Millionen-Jahre-Intervalls, in dem sich die Pflanzen zu Bäumen entwickelten und Wälder auf der Erde entstanden, nur 1,9-2,6 Mal über dem vorindustriellen Niveau lag.
Abb. 3: Zusammenfassung der Pflanzenevolution und der sich entwickelnden atmosphärischen Zusammensetzung im Vergleich zum Alter.
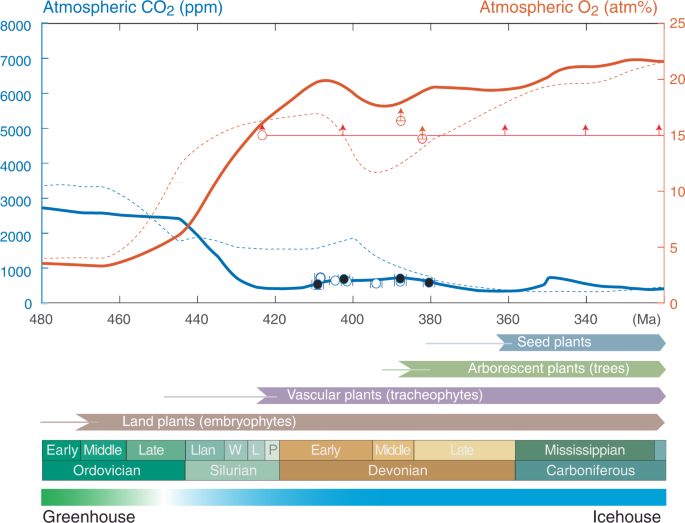
Atmosphärische pCO2-Beschränkungen von 410-380 Ma Lycophyten (Asteroxylon, Baragwanathia und Drepanophycus) mit Fehlerbalken, die kleiner als die Größe der Kreise sind. Gepaarte Spaltöffnungen und Isotopendaten vom selben Standort sind durch schwarze Kreise hervorgehoben. Der atmosphärische pO2-Wert wird durch Holzkohlebelege für Waldbrände seit ~423 Ma54, 55 (rote horizontale Linie) und fossile Wurzeln (rote Kreise)56 eingegrenzt, die nach kontrollierten Kalibrierungen im Labor56, 84 minimale pO2-Werte festlegen. Modellierte Entwicklung des atmosphärischen pCO2 (blaue Kurve) und des atmosphärischen pO2 (rote Kurve), abgeleitet durch Anpassung des COPSE Reloaded-Modells für die gekoppelten biogeochemischen Zyklen, so dass die kontinentale Verwitterungseffizienz früher vegetativer Ökosysteme mit der physikalischen Verwitterung von Feinstoffen in terrestrischen Ablagerungen46 und die Ausgasung aus der Erde mit dem kontinentalen Bogenvulkanismus skaliert. Das "COPSE reloaded"-Modell ist zum Vergleich dargestellt (dünne, gestrichelte Kurven)12. Für weitere Einzelheiten siehe ergänzende Informationen. Die Entstehung von Landpflanzen, Gefäßpflanzen, verzweigten Pflanzen mit tiefem Wurzelsystem und Samenpflanzen ist mit dünnen Linien dargestellt, die ihren Ursprung anhand von Schätzungen der molekularen Uhr darstellen, und mit dickeren Pfeilen, die fossile Vorkommen repräsentieren45. Llan Llandovery, W Wenlock, L Ludlow, P Pridoli. Die Unsicherheit der absoluten Alterszuweisungen ist biostratigraphisch definiert und beträgt etwa ±1,5 Ma.
Zwei Faktoren, die das Ausmaß der Isotopenfraktionierung des Blattkohlenstoffs beeinflussen können, sind Feuchtigkeit und atmosphärischer O2-Gehalt. Erstens regulieren die Pflanzen die stomatäre Leitfähigkeit herunter und erhöhen ihre Wassernutzungseffizienz in trockeneren Lebensräumen38. Lycophyten verfügen über eine passive Spaltöffnungssteuerung und können sich an trockenere Lebensräume anpassen, indem sie die Spaltöffnungsdichte und -größe minimieren und/oder mit einer geringeren CO2-Assimilationsrate überleben, was sich dann in einer geringeren Δ-Blattgröße ausdrücken würde27. Wir haben diese Anpassung experimentell mit H. squarrosa überprüft, die halb-epiphytisch (ergänzende Abb. 6) unter trockeneren Bedingungen (RH~60%) wuchs, was zu einem signifikant niedrigeren ΔBlatt (13,3 ± 0,2) als das ΔBlatt (20,1 ± 0,9) von Pflanzen führte, die unter optimaler Feuchtigkeit (RH~80%; ergänzende Tabelle 2) wuchsen. Dieser Effekt wird auch bei der Isotopenzusammensetzung natürlicher Populationen von C3-Pflanzen beobachtet, ist jedoch nur in Gebieten signifikant, in denen die durchschnittliche jährliche Niederschlagsmenge unter ~1000 mm/Jahr liegt (ergänzende Abb. 10)28. Im trockeneren Gewächshaus wiesen die H. squarrosa-Pflanzen eine geringere, wenn auch variablere Spaltöffnungsdichte (16,1 ± 4,7 vs. 20,1 ± 2,8 mm-2) und eine ähnliche Porenlänge (22,8 ± 1,0 µm vs. 21,6 ± 2,7 µm) auf. Obwohl wir nicht wissen, ob sich diese Lycophyten vollständig an die trockeneren Gewächshausbedingungen angepasst haben, ergibt die aus diesem Pflanzenmaterial abgeleitete pCO2-Schätzung eine Unterschätzung des pCO2-Niveaus im Gewächshaus um -128+79-67128-67+79 ppm, was durch ein etwas niedrigeres Verhältnis zwischen operativem und maximalem stomatären Leitwert (ζ) als bei Pflanzen erklärt werden könnte, die unter idealen (natürlichen) Bedingungen gewachsen sind (z. B. ζ von 0,14 vs. 0,20; siehe ergänzende Informationen für Details).
Um diesen Feuchtigkeitseffekt weiter zu bewerten, haben wir ein Atmosphäre-Ozean-Paläoklimamodell mittlerer Komplexität (CLIMBER-3α) verwendet, um das devonische Klima mit 500 ppm atmosphärischem CO2 zu bewerten, also am unteren Ende des Bereichs unserer neuen CO2-Schätzung. Das Modell wurde ausgeführt, um die Bedingungen, unter denen die frühen Lycophyten lebten, besser einzugrenzen (Einzelheiten siehe ergänzende Informationen) und zeigt einen gemäßigten Planeten mit mittleren tropischen Oberflächentemperaturen von 24,1-24,6 °C. Die Lycophytenflora in Australien, Deutschland und China wuchs im Monsungürtel mit hohen mittleren Jahresniederschlägen (MAR) von mehr als ~1000 mm/Jahr. Die Flora von Asteroxylon (Rhynie Chert, Schottland) und Drepanophycus aus Maine, USA, New Brunswick und Gaspé, Kanada, findet sich dagegen in den etwas trockeneren subtropischen Zonen. Aufgrund seiner groben räumlichen Auflösung kann unser Modell lokale Schwankungen der Niederschläge und der Luftfeuchtigkeit an den fossilen Fundorten nicht genau erfassen, aber die Modellergebnisse deuten auf ein Wachstum in feuchten oder relativ feuchten Umgebungen hin. Wichtig ist, dass wir keine Kopplung zwischen dem Δleaf oder dem vorhergesagten pCO2 der einzelnen Fossilfundorte und dem modellierten MAR, RH% oder dem Paläolatitudo sehen. Außerdem deuten die ΔBlattdaten nicht auf aride Wachstumsbedingungen hin. In Anbetracht dieser Daten gibt es keinen Hinweis darauf, dass die devonischen Lycophyten unter semi-ariden/ariden Bedingungen lebten oder dass der hier berichtete Paläo-pCO2-Proxy eine verzerrte Vorhersage in Abhängigkeit von einer Herabregulierung der stomatären Leitfähigkeit aufgrund des Wachstums in einer ariden Umgebung liefern sollte.
Zweitens kann atmosphärischer O2 auch die Kohlenstoff-Isotopenfraktionierung im Blatt beeinflussen, da die photosynthetische CO2-Fixierung mit der photosynthetischen O2-Fixierung des Rubisco-Enzyms konkurriert39. Dieser Effekt ist bei modernen Pflanzen, die bei subambientem CO2- und superambientem O2-Niveau wachsen, stärker ausgeprägt40. Die atmosphärischen pO2-Werte im Devon lagen wahrscheinlich bei ~15 bis 20 atm%12 und damit nicht viel niedriger als heute, so dass der O2-Effekt auf unsere Paläo-pCO2-Schätzungen wahrscheinlich gering ist. Kontrollierte Wachstumsexperimente mit Gefäßpflanzen, darunter eine Lycophytenart (Selaginella kraussiana), zeigen eine kleine positive Verschiebung des ΔBlattes (0,5 ± 0,4), wenn es bei einem O2-Gehalt unterhalb der Umgebungsluft (16 atm%) wächst40. Dieser Effekt wurde der O2-Empfindlichkeit des photorespiratorischen Kompensationspunktes zugeschrieben41 und hat im Vergleich zur O2-unempfindlichen Parametrisierung des mechanistischen Proxys eine vernachlässigbare Auswirkung auf vergangene atmosphärische CO2-Schätzungen (ergänzende Tabelle 4).
Wichtig ist, dass unsere neuen und präziseren CO2-Vorhersagen radikal niedriger sind als die zuvor gemeldeten Mittelwerte, aber sie liegen innerhalb des Unsicherheitsbereichs anderer pCO2-Proxys, einschließlich der pedogenen Karbonataufzeichnungen, wenn man bedenkt, dass die devonischen Böden wohl weniger produktiv waren, mit geringeren Bodenatmungsraten17 und niedrigeren CO2-Konzentrationen als moderne Böden (ergänzende Abb. 14).
Die Umgestaltung des Blattgasaustauschmodells zur Lösung der CO2-Assimilationsrate (A0) ermöglicht es uns, die ökophysiologische Leistung früher Landpflanzen zu untersuchen, die keine lebenden Verwandten haben. Dieser Ansatz ermöglicht es uns, die sehr geringe Stomatendichte dieser Pflanzen zu kontextualisieren. Die Modellergebnisse deuten darauf hin, dass diese Pflanzen wesentlich geringere CO2-Assimilationsraten hatten als die zeitgenössischen Lycophyten (A0 < 3,7 μmol m-2 s-1); Aglaophyton (0,40 ± 0,10 μmol m-2 s-1), Rhynia (0. 62 ± 0,13 μmol m-2 s-1), Horneophyton (0,84 ± 0,21 μmol m-2 s-1), Sawdonia (0,89 ± 0,04 μmol m-2 s-1) und Nothia (1,27 ± 0,29 μmol m-2 s-1) (Supplementary Data 1). Es ist daher plausibel, dass die niedrigen Assimilationsraten in dieser Pflanzengruppe ein Faktor für ihre eventuelle Verdrängung sein könnten. Die Berechnung der niedrigen Assimilationsraten legt nahe, dass der Sporophyt physiologisch an den Gametophyten gebunden sein könnte, was auf eine gewisse Matrotrophie hindeutet, wie sie für einige Cooksonia-Arten vermutet wird42.
CO2 in der Atmosphäre und frühe Aufforstung
Die atmosphärische pCO2-Kurve (Abb. 3) gibt Aufschluss darüber, wie sich die frühe Geschichte der Landpflanzen und ihre Besiedlung der Erdoberfläche auf das Klimasystem der Erde auswirkten. Die Entstehung der Bäume im mittleren Devon (393-383 Ma)1 stellt einen bedeutenden evolutionären Wandel dar, bei dem sich die Arboreszenz unabhängig voneinander in drei verschiedenen Pflanzenstämmen (Lycopsiden, Sphenopsiden, Farne) entwickelte. Progymnosperme Bäume, die bis zu ~30 m hoch werden, diversifizierten sich und verbreiteten sich von paläoäquatorialen zu paläoborealen Breiten und von saisonal trockenen zu feuchten Lebensräumen1,43. Im Gegensatz zur früheren, von Lycophyten dominierten Flora, die typischerweise ~0,1 m hoch war und deren Rhizoide nur wenige cm in den Boden eindrangen, hatten die sich entwickelnden Bäume sowohl tiefere Wurzeln zur Verankerung als auch ein hochentwickeltes Wurzelsystem, das dem moderner Samenpflanzen ähnelte1. Dennoch blieb der atmosphärische CO2-Gehalt recht niedrig und könnte während der Entstehung der Wälder im Devon höchstens um einige hundert ppm gesunken sein17.
Das absolute Ausmaß des frühpaläozoischen pCO2-Rückgangs ist immer noch nicht genau bekannt, und es besteht ein Konflikt zwischen einigen niedrigen pCO2-Schätzungen, die aus den Aufzeichnungen von marinen Phytanen aus dem mittleren Ordovizium stammen, und den hohen pCO2-Schätzungen aus pedogenem Goethit18. Wenn wir davon ausgehen, dass der pCO2-Wert in der spätordovizischen Atmosphäre höher war als heute, zumindest außerhalb der eiszeitlichen Maxima (ergänzende Abb. 12), deuten unsere neuen Ergebnisse auf einen dramatischen und abrupten Rückgang von ~9 (ungebundenen) PIAL auf ~1,9 ± 0,3 PIAL hin, der in einem relativ kurzen Zeitintervall von ~445-410 Ma vor dem Auftreten von Wäldern stattfand. Gleichzeitig breiteten sich strauchartige Gefäßpflanzen auf den Kontinenten aus (z. B. die Eophytenflora44), und eine dramatische Veränderung des physikalischen Verwitterungsregimes, die durch die sich entwickelnden terrestrischen Ökosysteme angetrieben wurde, ist durch die Speicherung von feinkörnigem Sediment in kontinentalen Ablagerungen belegt45,46,47. Wir vermuten, dass die früheste vaskuläre Vegetation die Exposition einer größeren mineralischen Oberfläche gegenüber Verwitterungsflüssigkeiten förderte und die globale Silikatverwitterung auf den Kontinenten verstärkte (weit mehr, als dies spätere, tief verwurzelte Ökosysteme tun konnten), was auf einen größeren Nährstoffverlust aus weniger entwickelten Böden zurückzuführen ist und somit einen höheren Verwitterungsbedarf erzwang.
Um die Auswirkungen der Etablierung früher Gefäßpflanzen-Ökosysteme zu simulieren, haben wir ein dynamisches Modell (COPSE Reloaded) für die gekoppelten biogeochemischen Kreisläufe verwendet, um die atmosphärischen pCO2- und pO2-Verläufe vorherzusagen (siehe Methoden). Unser Modell sagt einen massiven Rückgang des atmosphärischen CO2 von ~2500 ppm auf ~500 ppm in nur ~30 Myr als Reaktion auf die verstärkte Silikatverwitterung durch frühe Gefäßpflanzen voraus (Abb. 3). Um ein solches Szenario zu veranschaulichen, haben wir mehrere Antriebsfunktionen im neuesten COPSE-Modell12,48,49 aktualisiert (Abschnitt S5; Ergänzende Abbildungen 19-21). Der Verwitterungsantrieb (W) wurde so angepasst, dass er proportional zum pflanzeninduzierten Effekt auf die Schlammgestein-Retention in kontinentalen Ablagerungen skaliert und auf den karbonischen Durchschnitt normiert wird46. Dies ist gerechtfertigt, da die Mineraloberfläche ein Schlüsselfaktor ist, der die Mineralauflösung während der chemischen Verwitterung erleichtert. Außerdem haben wir die Pflanzenevolution in Übereinstimmung mit der Ausbreitung von Gefäßpflanzen anstelle von Nicht-Gefäßpflanzen in früheren COPSE-Modellen hochskaliert, um sowohl eine umfangreichere Pflanzenabdeckung50,51 zu erfassen als auch die selektive P-Verwitterung zu berücksichtigen, um einen größeren Verwitterungsbedarf früher Gefäßpflanzen mit primitiven wurzelähnlichen Systemen zu imitieren49,50,51. Schließlich wurden die vulkanischen Ausgasungsraten im Unter- und Mitteldevon so angepasst, dass der aus dem Erdinneren stammende CO2-Fluss mit den subduzierten Karbonatplattformen skaliert und nicht mit den globalen Ausbreitungsraten des Meeresbodens, wie in früheren Modellen angenommen wurde11,52. Dazu wurde eine lineare Beziehung zwischen der Ausgasungsrate, dem Vulkanismus der Kontinentalbögen und der relativen Häufigkeit junger und älterer Körner, die aus dem Bogen stammen, in den Sedimentablagerungen angenommen53. Die Ausgasungsrate vom späten Silur bis zum mittleren Devon (~430-370 Ma) beträgt dann das ~1,0- bis 1,2-fache des heutigen Niveaus (im Gegensatz zu ~1,5 in früheren Modellen), was charakteristisch für das Alter ist, in dem sich Superkontinente zusammensetzen.
Das überarbeitete Modell sagt gleichzeitig voraus, dass die sich entwickelnden vaskulären Ökosysteme auch einen Anstieg des atmosphärischen pO2 verursachten, der mit mehreren Linien geochemischer Beweise für die Sauerstoffanreicherung der Erde12 übereinstimmt, einschließlich der Beweise für Wildfeuer, die atmosphärische O2-Werte über 15 atm% seit ~420 Ma54,55 und fossile Wurzeln56 belegen (Abb. 3). Frühere Modelle haben einen zweistufigen Übergang mit relativ hohem pCO2 und hohem pO2 im Silur12 vorgeschlagen, aber unsere neuen Daten und unser Modell bieten eine einfachere Lösung, bei der ein synchroner Anstieg des atmosphärischen O2 und ein Rückgang des CO2 auf nahezu moderne Bedingungen im selben Ereignis stattfanden.
Letztendlich wird die Zusammensetzung der Erdatmosphäre durch ein Zusammenspiel von biologischen und geologischen Prozessen bestimmt und dadurch, wie Landpflanzen und ihre Wurzelsymbionten die physikalischen und chemischen Verwitterungsprozesse an Land beeinflussen45,57. Der Unterschied im atmosphärischen CO2 durch die Aufforstung nach der Besiedlung mit Gefäßen betrug höchstens ein paar hundert ppm58. Die globale Abkühlung des Klimas hängt also nicht so sehr mit der Aufforstung zusammen59 , sondern vielmehr damit, wie die terrestrische Biosphäre Nährstoffe aus ihrem planetarischen Substrat extrahiert und erhält.
Das Klima des mittleren Paläozoikums neu betrachtet
Das Klima im Devon wurde bisher meist als warmes Treibhaus beschrieben, das mit der polaren Vergletscherung im Spätdevon-Karbon in einen kälteren Zustand überging10,15. Bei nur 500 ppm CO2 sagt unser frühdevonisches Paläoklimamodell jedoch ein mildes Klima mit globalen Jahresmitteltemperaturen von 14,8-15,7 °C für verschiedene Orbitalkonfigurationen und einem starken latitudinalen Temperaturgradienten voraus, der sich nicht allzu sehr von dem heutigen unterscheidet. Die mittleren tropischen Oberflächentemperaturen liegen bei 24,1-24,6 °C. Die vorhergesagten Meeresoberflächentemperaturen im tropischen Gondwana, in Südchina, West-Laurentien und Europa stimmen mit früh- und mitteldevonischen Paläotemperaturen überein, die auf der Sauerstoffisotopenzusammensetzung von phosphatischen Conodonten und gut erhaltenen kalzitischen Brachiopoden basieren (ergänzende Daten 2)60,61. Unser Paläoklimamodell mit 500 ppm atmosphärischem CO2 sagt deutlich niedrigere Temperaturen in der Subtropen- und Temperaturzone voraus, für die es derzeit keine präzisen Paläotemperaturschätzungen gibt (siehe ergänzende Informationen, Abschnitt S4.2 für weitere Diskussionen)62. Polares Meereis und Schneefall auf Gondwana werden für die Winter vorhergesagt (Abb. 4, ergänzende Abb. 14-16). CLIMBER-3α ist nicht mit einem dynamischen Eisschildmodell gekoppelt, so dass die Ausdehnung und das Fortbestehen von Eisschilden indirekt durch die Ausdehnung von Schnee und Meereis bestimmt wird. Wir stellen fest, dass das Erdsystem klimatisch stabil war und dass eine Kaskade in eine schneeballartige Vergletscherung aufgrund der Eis-Albedo-Rückkopplung bei diesem niedrigen CO2-Niveau selbst unter der schwächeren Sonneneinstrahlung höchst unwahrscheinlich ist, ein Ergebnis, das auch von Modellsimulationen über das frühe Devon63,64,65 unterstützt wird. Daher deuten unsere Ergebnisse darauf hin, dass sich das Klima der Erde in einem Eishaus-Zustand mit teilweiser Vergletscherung auf dem Südpolkontinent Gondwana oberhalb von 60-80°S Paläolatitude befand (Abb. 4). Ausgehend von modernen Paläoklimamodellen14,66,67 schlagen wir vor, die ursprüngliche Vermutung10 , dass mehrere Tausend ppm CO2 in der Atmosphäre notwendig waren, um die ~3% schwächere paläozoische Sonne zu kompensieren, zugunsten einer viel höheren Klimasensitivität von CO2 aufzugeben, wie sie von den neuen Paläo-CO2-Proxydaten3 unterstützt wird.
Abb. 4: Jährliche mittlere Oberflächenlufttemperatur im frühen Devon (415 Ma) für einen atmosphärischen pCO2 von 500 ppmv, modelliert mit dem gekoppelten Klimamodell CLIMBER-3α.
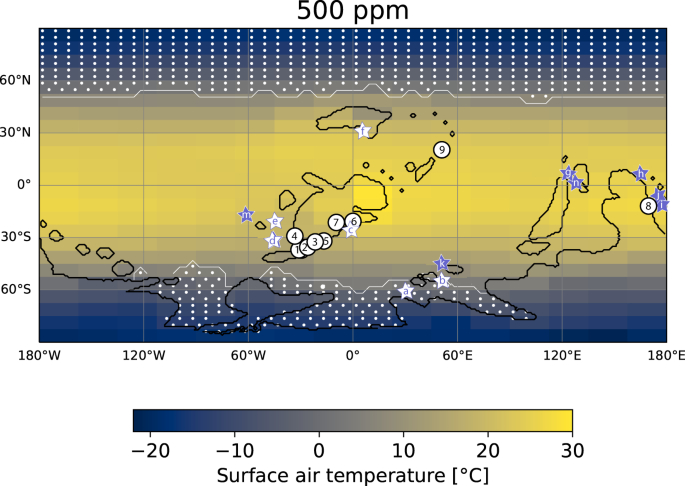
Es wird die paläogeographische Konfiguration von Schottland bei 415 Ma83 verwendet, vorindustrielle Werte anderer Treibhausgase und idealisierte Orbitalparameter werden angenommen (kreisförmige Umlaufbahn, Schiefe von 23,5°; siehe ergänzende Abb. 17 für zwei verschiedene Orbitalzustände). Einzelheiten zu den modellierten Niederschlägen und der Luftfeuchtigkeit finden sich im Anhang. Neun Orte sind markiert, an denen die untersuchte fossile Lycophytenflora in 13 verschiedenen Aufschlüssen gefunden wurde: 1 - Green County, NY, USA und Schoharia Co, NY, USA; 2 - Traveller Mountain, ME, USA; 3 - North Shore und Seal Rock, Gaspe Qbc, Kanada; 4 - Abitibi River, Ontario, Kanada; 5 - Dalhousie, New Brunswick, Kanada (in der Nähe von Maple Green); 6 - Munchshecke, Siegburg und Moseltal, Deutschland; 7 - Rhynie Chert, Aberdeenshire, Schottland; 8- Yea District, Victoria, Australien; 9 - Xinjan, China. Sechs Fundorte mit Paläo-SST-Daten von kalzitischen Brachiopoden sind mit weißen Sternen gekennzeichnet: a - Abschnitt Madène el Mrakib, Anti-Atlas-Gebirge, Marokko; b - Colle-Gebiet, Kantabrisches Gebirge, Spanien; c - Eifel, Deutschland; d - Buffalo Quarry, Iowa, USA; e- Mafeking Quarry, Manitoba, Kanada; f- Stadt Gur'evsk, Altai-Gebirge/Salair-Gebiet, Sibirien. Sieben Orte mit Paläo-SST-Daten aus phosphatischen Conodonten sind mit lila Sternen markiert: g - Changputang, SE Yunnan, China und Nayi, Guangxi, China; h - Broken River, Queensland, Australien; i - Buchan, Boola Quarry, Tyers Quarry, Wellington, Victoria, Australien; j - Mungallala und Windellama, Wellington New South Wales, Australien; k - Barrandov und Na Skrabku, Prager Becken, Tschechische Republik; m - Nevada, USA; n - CaiZiyan, Guangxi, China. Phosphatische SST-Daten aus d - Glory Quarry, Iowa, USA, und b - Puech de la Suque, Frankreich, sind zusammen mit den kalzitischen SST-Daten von nahe gelegenen Aufschlüssen dargestellt.
Direkte Beweise für die Vergletscherung finden sich in spätdevonisch-karbonischen Ablagerungen68 , sind aber in unter- und mitteldevonischen Schichten selten. Indirekte Beweise für eustatische Meeresspiegelveränderungen finden sich in den unterdevonischen Sedimentabfolgen aus Nordafrika, die möglicherweise einen gletscherbedingten Ursprung haben könnten69. Des Weiteren sind sauerstoffisotopische Paläotemperaturaufzeichnungen60 aus den flachen tropischen und subtropischen Ozeanen mit den von uns vorhergesagten Paläoklimadaten vereinbar, zeigen aber auch erhebliche lokale Temperaturschwankungen61. Außerdem finden sich devonische Pflanzen meist in niedrigeren bis mittleren Paläolatritäten (~45°S), wo der Kontinent vermutlich dauerhaft eisfrei ist. Eine Ausnahme könnte die frühe lockhovianische Cooksonia-reiche Flora aus dem Paraná-Becken in Brasilien sein, das in hohen Paläolatitudes (~70°S) liegt70. Doch selbst bei 500 ppm CO2 zeigt sich, dass die Schneedecke auf Gondwana nicht immer ganzjährig war (ergänzende Abb. 15). Obwohl die Paraná-Flora deutlich vor unserer Aufzeichnung (~419 Ma) liegt, als die atmosphärischen CO2-Werte vermutlich höher waren, könnten Pflanzen auch in hohen Breitengraden gewachsen sein, entweder während wärmerer südlicher Sommer oder weil das Klima aus anderen Gründen als hohem CO2 wärmer war (z. B. höheres CH4, N2O).
Zusammenfassend lässt sich sagen, dass der revidierte atmosphärische CO2-Datensatz mit allen paläoklimatischen Vorgaben aus der geologischen Aufzeichnung, mit globalen Paläoklimamodellen und mit globalen Modellen für die gekoppelten biogeochemischen Kreisläufe vereinbar ist, die einen größeren Verwitterungsbedarf der frühesten terrestrischen Flora berücksichtigen und bei denen die frühe Aufforstung nur eine geringe Rolle für die globale CO2-Sequestrierung spielte.
Methoden
Proben
Kohlenstoffisotope, Spaltöffnungsdichte und Spaltöffnungsgröße von fossilen Blättern wurden aus der Literatur zusammengestellt (siehe ergänzende Informationen), zusammen mit neuen Daten aus der Baragwanathia-Flora in Victoria, Australien. Pflanzenmakrofossilien aus der Fossiliensammlung des Museums Victoria wurden ursprünglich von Isabel Cookson in Mt Pleasant Rd, Victoria, Australien, gesammelt71. Die Fossilien sind als Inkrustationen in feinkörnigem Sandstein erhalten, der in einem 130 m dicken stratigraphischen Abschnitt aus überwiegend Schluffstein und Schiefer liegt (ergänzende Abb. 1). Sie werden als gelegentliche Ausbrüche hochenergetischer Turbidite interpretiert, die allochthone Fossilien aus flacheren Gewässern in ein sehr energiearmes marines Ablagerungsmilieu tragen72. Die Indexfossilien (Uncinatograptus sp. cf. U. thomasi und Nowakia sp. ex gr. N. acuaria) grenzen die Flora auf das Pragian oder früheste Emsian ein, was laut GTS202073 ~409,1 ± 1,5 Ma entspricht.
Kleine Fossilfragmente, die in vier Exemplaren (#15154, #15173, #15174, #15183) gefunden wurden, die auch größere Fragmente von Baragwanathia longifolia und Zosterophyllum australianum enthalten, wurden aufgrund der Erhaltung der schwarzen organischen Substanz, die in den braunen mineralisierten Fossilien enthalten ist, für Analysen ausgewählt (ergänzende Abb. 2-4). Von jedem Fragment (insgesamt 37) wurden 0,05-7,34 mg Material mit einem Skalpell oder einem 0,8 mm Dremel-Bohrer entnommen. Es wurde nur Material extrahiert, das sichtbar schwarze organische Substanz enthielt.
Kohlenstoff-Isotopenanalysen
Die Kohlenstoff-Isotopenanalysen wurden im GAPP-Labor der Syracuse University mit einem automatisierten "nano-EA"-System durchgeführt, das an das in Polissar et al.74 beschriebene System angepasst wurde. Das Nano-EA-System der Syracuse University besteht aus einem Elementar Isotope Cube Elementaranalysator, der über einen Isoprime Trace Gas Analyzer mit einem Isoprime 100 Massenspektrometer für stabile Isotope mit kontinuierlichem Fluss verbunden ist. Obwohl das Vorhandensein von Karbonat weder vermutet noch bei der Untersuchung der Pulverproben mit 6 N HCl unter einem Binokular beobachtet wurde, beschlossen wir, die Probenmaterialien vor der Analyse zu dekarbonisieren, um sicherzustellen, dass sie karbonatfrei sind. Die Pulverproben wurden 24 Stunden lang in einer evakuierten Glasglocke in einem Glasfläschchen in Gegenwart von reiner Salzsäure begast und anschließend im Ofen bei 40 °C getrocknet. Für die Isotopenanalyse wurde das Probenmaterial in Silber- oder Quarzbecher (6 ×6 mm; EA Consumables) gefüllt und in eine kleine Menge Quarzwolle eingebettet, um die Rückhaltung des Probenmaterials im Becher sicherzustellen. Die Becher und die Quarzwolle wurden 8 Stunden lang bei 480 °C verascht. Das Probenmaterial wurde in einer Isolierkabine mit laminarer Strömung der Klasse 100 und HEPA-gefilterter Luft in die Becher gefüllt, um das Risiko einer Partikelkontamination zu minimieren.
Während der Isotopenanalyse wurden die Probenbecher evakuiert und mit Helium gespült, bevor sie in den EA eingeführt wurden. Die Reaktionsbedingungen waren wie folgt: Die Temperaturen des Oxidations- und des Reduktionsreaktors betrugen 1100 °C bzw. 650 °C; der Helium-Trägergasfluss betrug 158 ml/min und der O2-Puls war auf 45 Sekunden eingestellt. Das bei der Verbrennung der Probe entstehende Kohlendioxid wurde im EA in einer Molekularsiebfalle aufgefangen. Nach dem Durchgang des N2-Peaks wurde die primäre EA-Falle erhitzt und das Kohlendioxid in eine sekundäre, mit Kieselgel gefüllte Kryofalle abgegeben, die in flüssigen Stickstoff getaucht war. Die Einfangdauer wurde anhand der Daten des EA-Wärmeleitfähigkeitsdetektors kalibriert, um eine vollständige Erfassung des CO2-Peaks sicherzustellen. Nach der CO2-Sammlung wurde der Gasfluss der Kryofalle über ein automatisiertes Vici Valco 6-Port-Ventil auf ein He-Trägergas mit geringerem Durchfluss (~1 mL/min) umgeschaltet. Die Falle wurde erwärmt, und das Probengas wurde über eine Agilent CarboBond-Kapillarchromatographiesäule (25 m x 0,53 mm x 5 µm) an das IRMS abgegeben. Die resultierenden Kohlenstoff-Isotopen-Rohdaten wurden durch direkte Leerwert-Subtraktion korrigiert und mit den internationalen Referenzmaterialien NIST 1547-Peach Leaves (δ13C = 26,0 ± 0,2) und IAEA C6-Sucrose (δ13C = -10,45 ± 0,03), die als Feststoffe verwendet werden, auf die VPDB-Skala normiert75. Die Referenzmaterialien USGS 61-Koffein (-35,05) und USGS 62-Koffein (-14,79) sind in UV-behandeltem MilliQ-Wasser gelöst, werden in bekannten Mengen dosiert und haben eine langfristige Laborreproduzierbarkeit von ±0,47 für USGS 62 und ±0,39 für USGS 61 über einen Bereich von 20-90 Nanomol. Die Reproduzierbarkeit der δ13C-Werte von Referenzmaterialien mit einem Kohlenstoffgehalt von mehr als 50 Nanomol beträgt ±0,3 (1 sd) und entspricht den zuvor berichteten Werten74.
TOF-SIMS-Analysen
Die Morphologie und die chemische Zusammensetzung der Fragmente wurden zerstörungsfrei mittels SEM und TOF-SIMS charakterisiert, um das Vorhandensein von organischem Gewebe zu bestätigen, das sich gut für die Kohlenstoffisotopenanalyse eignet. Mit Hilfe der Flugzeit-Sekundärionen-Massenspektrometrie (TOF-SIMS) wurden halbquantitative Karten der Elementzusammensetzung der Probe #15153 erstellt (siehe ergänzende Abb. 5).
Bei dieser Technik wird die Oberfläche mit Bi-Ionen beschossen, die eine Kollisionskaskade in den obersten Atomschichten der Probe (~10 nm) auslösen. Dadurch werden sekundäre Ionen freigesetzt, die in einem elektrischen Feld beschleunigt werden und deren Flugzeit zum Detektor im Vakuum von ihrer Masse und der Probentiefe abhängt76,77,78. Ergänzende Abb. 5 zeigt an organische Stoffe gebundene Elemente (einschließlich C, N, P) in der Probe, die polyatomar geladene Spezies wie CN- und CNO- erzeugen, wenn sie aus der gleichen Probentiefe emittiert werden. Die Korrelationen der organisch gebundenen Elemente ermöglichen es uns, das Vorhandensein von organischem Kohlenstoff von anorganischen Phasen (z. B. Karbonatmineralen) in der Probe zu unterscheiden.
Modellierung des Paläoklimas
Das relativ schnelle gekoppelte Erdsystemmodell mittlerer Komplexität CLIMBER-3α wurde verwendet, um das Klima im Devon mit einem atmosphärischen CO2-Gehalt von 500 ppm zu simulieren. CLIMBER-3α umfasst eine modifizierte Version des Ozeanzirkulationsmodells (MOM379,80) mit einer horizontalen Auflösung von 3,75° x 3,75° und 24 vertikalen Ebenen, ein dynamisches/thermodynamisches Meereismodell81 mit der gleichen Auflösung und ein schnelles atmosphärisches Modell82 mit einer Auflösung von 22,5° in der Länge und 7,5° in der Breite. Das Modell modelliert nicht explizit das Wachstum von Eisschilden auf den Kontinenten, aber die Schneedecke auf den Kontinenten wird berücksichtigt. Das Modell wurde für unterdevonische Randbedingungen (415 Ma) in Bezug auf Kontinentalkonfiguration, Sonneneinstrahlung und Vegetationsbedeckung durchgeführt83. Auf der Grundlage früherer Ergebnisse67 wurden drei verschiedene Orbitalkonfigurationen untersucht: die Standardkonfiguration (Schiefe 23,5°, Exzentrizität e = 0) sowie kalte (Schiefe 22,0°, Exzentrizität e = 0) und warme (Schiefe 24,5°, Exzentrizität e = 0,069, Präzessionswinkel 0°) Orbitalkonfigurationen (ergänzende Abbildung 17). Eine Sensitivitätsanalyse unter Berücksichtigung der saisonalen Oberflächenlufttemperaturen und der Meereisverteilung für diese unterschiedlichen Einstrahlungsmuster ist in der ergänzenden Tabelle 6 dargestellt. Zur Bestimmung der Klimavariablen an bestimmten Proxy-Standorten (siehe ergänzende Daten 2) wurden die simulierten Werte auf einem 1°-mal-1°-Gitter bilinear interpoliert und unter Verwendung von Koordinaten ausgewertet, die mit GPlates aus den heutigen Werten transformiert wurden.
Langfristige globale biogeochemische Modellierung
Wir haben das Modell der Kohlenstoff-Sauerstoff-Phosphor-Schwefel-Evolution (COPSE) verwendet, um die Entwicklung des atmosphärischen pCO2, pO2 und der Ozeanzusammensetzung im Phanerozoikum (550 Ma bis heute) vorherzusagen. Dieser Vorwärtsmodellierungsansatz ermöglicht die Prüfung von Hypothesen über mechanistische Ursache-Wirkungs-Beziehungen im Erdsystem. Eine Reihe von gekoppelten Differentialgleichungen, die die dynamische Entwicklung der C-, O-, P- und S-Zyklen beschreiben, wurden mit einem eingebauten Löser für "starre" gewöhnliche Differentialgleichungssysteme in Matlab® mit variablen Zeitschritten gelöst. Die neueste Version des Modells (COPSE Reloaded, bezeichnet als "CR"; ergänzende Abbildungen 17-18) wurde angepasst und modifiziert48,49. Als Input ließen wir das C/P-Verhältnis der vergrabenen terrestrischen Biomasse mit der Besiedlung des Landes durch nicht-vaskuläre Pflanzen wie in CR ansteigen und passten die Einflüsse auf die Pflanzenverwitterung (W), die Pflanzenevolution (E), die selektive P-Verwitterung (F) und die vulkanische Ausgasung (D) so an, dass sie gleichzeitig die Auswirkungen flacher vaskulärer Ökosysteme auf die Verwitterungsprozesse berücksichtigen und Ergebnisse liefern, die mit den Paläoaufzeichnungen übereinstimmen. Abbildung 3 zeigt die atmosphärischen pO2- und pCO2-Trajektorien, die von der überarbeiteten Version von CR vorhergesagt wurden, mit den in der ergänzenden Abbildung 21 dargestellten Antriebsfaktoren. Weitere Einzelheiten zur überarbeiteten COPSE-Modellierung finden sich im ergänzenden Textabschnitt S5.
Verfügbarkeit von Daten
Alle Daten sind im Haupttext oder in den ergänzenden Materialien verfügbar.
Verfügbarkeit des Codes
Die in dieser Analyse entwickelten Modelle werden im elektronischen Forschungsdatenarchiv (ERD) der Universität Kopenhagen zur Verfügung gestellt. https://doi.org/10.17894/ucph.214a6434-b7eb-4e62-aaac-afefc1247da4. Der Quellcode für das in dieser Studie verwendete Paläoklimamodell CLIMBER ist im Potsdam-Institut für Klimafolgenforschung archiviert und wird auf Anfrage zur Verfügung gestellt. Die Postprocessing-Skripte, die zur Analyse der CLIMBER-Daten und zur Erstellung der in der Studie präsentierten Paläoklimamodell-Zahlen verwendet wurden, sind auf ERDA gespeichert.
Referenzen
- Stein, W. E. et al. Mid-devonian archaeopteris roots signal revolutionary change in earliest fossil forests. Curr. Biol. 30, 421431.e2 (2020).Article CAS Google Scholar
- Foster, G. L., Royer, D. L. & Lunt, D. J. Future climate forcing potentially without precedent in the last 420 million years. Nat. Commun. 8, 14845 (2017).Article ADS CAS Google Scholar
- Berner, R. A. Paleozoic atmospheric CO2: importance of solar radiation and plant evolution. Science 261, 6870 (1993).Article ADS CAS Google Scholar
- Walker, J. C. G., Hays, P. B. & Kasting, J. F. A negative feedback mechanism for the long-term stabilization of Earths surface temperature. J. Geophys. Res. 86, 9776 (1981).Article ADS CAS Google Scholar
- DAntonio, M., Ibarra, D. E. & Boyce, C. K. Land plant evolution decreased, rather than increased, weathering rates. Geology https://doi.org/10.1130/G46776.1 (2019).Article Google Scholar
- Caves, J. K., Jost, A. B., Lau, K. V. & Maher, K. Cenozoic carbon cycle imbalances and a variable weathering feedback. Earth Planet. Sci. Lett. 450, 152163 (2016).Article ADS CAS Google Scholar
- Maher, K. & Chamberlain, C. P. Hydrologic regulation of chemical weathering and the geologic carbon cycle. Science 343, 15021504 (2014).Article ADS CAS Google Scholar
- Goddéris, Y. et al. Onset and ending of the late Palaeozoic ice age triggered by tectonically paced rock weathering. Nat. Geosci. 10, 382386 (2017).Article ADS Google Scholar
- Ibarra, D. E. et al. Modeling the consequences of land plant evolution on silicate weathering. Am. J. Sci. 319, 143 (2019).Article ADS CAS Google Scholar
- Berner, R. A. Atmospheric carbon dioxide levels over phanerozoic time. Science 249, 13821386 (1990).Article ADS CAS Google Scholar
- Berner, R. A. GEOCARBSULF. A combined model for Phanerozoic atmospheric O2 and CO2. Geochimica et Cosmochimica Acta 70, 56535664, (2006).
- Lenton, T. M. et al. Earliest land plants created modern levels of atmospheric oxygen. Proc. Natl. Acad. Sci. USA 113, 97049709 (2016).Article ADS CAS Google Scholar
- Ghienne, J.-F. et al. A Cenozoic-style scenario for the end-Ordovician glaciation. Nat. Commun. 5, 4485 (2014).Article ADS CAS Google Scholar
- Pohl, A. et al. Glacial onset predated Late Ordovician climate cooling. Paleoceanography 31, 800821 (2016).Article ADS Google Scholar
- Royer, D. L. Stomatal density and stomatal index as indicators of paleoatmospheric CO2 concentration. Rev. Palaeobot. Palynol. 114, 128 (2001).Article Google Scholar
- Mora, C. I., Driese, S. G. & Colarusso, L. A.Middleto Late Paleozoic Atmospheric CO$_2$ Levels from Soil Carbonate and Organic Matter. Sci. New Ser. 271, 11051107 (1996).CAS Google Scholar
- Ekart, D. D. A 400 million year carbon isotope record of pedogenic carbonate; implications for paleoatomospheric carbon dioxide. Am. J. Sci. 299, 805827 (1999).Article ADS CAS Google Scholar
- Witkowski, C. R., Weijers, J. W. H., Blais, B., Schouten, S. & Sinninghe Damsté, J. S. Molecular fossils from phytoplankton reveal secular P co 2 trend over the Phanerozoic. Sci. Adv. 4, eaat4556 (2018).Article ADS CAS Google Scholar
- Yapp, C. J. & Poths, H. Ancient atmospheric CO2 pressures inferred from natural goethites. Nature 355, 342344 (1992).Article ADS CAS Google Scholar
- McElwain, J. & Chaloner, W. Stomatal density and index of fossil plants track atmospheric carbon dioxide in the palaeozoic. Ann. Bot. 76, 389395 (1995).Article Google Scholar
- Yapp, C. J. A possible goethite-iron(III) carbonate solid solution and the determination of CO2 partial pressures in low-temperature geologic systems. Chem. Geol. 64, 259268 (1987).Article ADS CAS Google Scholar
- Russell, J. D., Paterson, E., Fraser, A. R. & Farmer, V. C. Adsorption of carbon dioxide on goethite (α-FeOOH) surfaces, and its implications for anion adsorption. J. Chem. Soc. Faraday Trans. 1: Phys. Chem. Condens. Phases 71, 1623 (1975).Article CAS Google Scholar
- Breecker, D. O., Sharp, Z. D. & McFadden, L. D. Atmospheric CO2 concentrations during ancient greenhouse climates were similar to those predicted for A.D. 2100. Proc. Natl. Acad. Sci. 107, 576580 (2010).Article ADS CAS Google Scholar
- Edwards, D. Stomata in early land plants: an anatomical and ecophysiological approach. J. Exp. Bot. 49, 255278 (1998).Article Google Scholar
- Witkowski, C. R., van der Meer, M. T. J., Smit, N. T., Sinninghe Damsté, J. S. & Schouten, S. Testing algal-based pCO2 proxies at a modern CO2 seep (Vulcano, Italy). Sci. Rep. 10, 10508 (2020).Article ADS CAS Google Scholar
- Freeman, K. H. & Hayes, J. M. Fractionation of carbon isotopes by phytoplankton and estimates of ancient CO 2 levels. Glob. Biogeochem. Cycles 6, 185198 (1992).Article ADS CAS Google Scholar
- Franks, P. J. et al. New constraints on atmospheric CO2 concentration for the Phanerozoic. Geophys. Res. Lett. 41, 46854694 (2014).Article ADS CAS Google Scholar
- Diefendorf, A. F., Mueller, K. E., Wing, S. L., Koch, P. L. & Freeman, K. H. Global patterns in leaf 13C discrimination and implications for studies of past and future climate. Proc. Natl Acad. Sci. 107, 57385743 (2010).Article ADS CAS Google Scholar
- Schubert, B. A. & Jahren, A. H. Global increase in plant carbon isotope fractionation following the Last Glacial Maximum caused by increase in atmospheric pCO2. Geology 43, 435438 (2015).Article ADS CAS Google Scholar
- Schubert, B. A. & Jahren, A. H. The effect of atmospheric CO2 concentration on carbon isotope fractionation in C3 land plants. Geochimica et. Cosmochimica Acta 96, 2943 (2012).Article ADS CAS Google Scholar
- Cui, Y. & Schubert, B. A. A 23 m.y. record of low atmospheric CO2. Geology 48, 888892 (2020).Article ADS CAS Google Scholar
- Diefendorf, A. F., Leslie, A. B. & Wing, S. L. Leaf wax composition and carbon isotopes vary among major conifer groups. Geochimica et. Cosmochimica Acta 170, 145156 (2015).Article ADS CAS Google Scholar
- Lomax, B. H., Lake, J. A., Leng, M. J. & Jardine, P. E. An experimental evaluation of the use of Δ13C as a proxy for palaeoatmospheric CO2. Geochimica et. Cosmochimica Acta 247, 162174 (2019).Article ADS CAS Google Scholar
- Stein, R. A., Sheldon, N. D. & Smith, S. Rapid response to anthropogenic climate change by Thuja occidentalis: implications for past climate reconstructions and future climate predictions. PeerJ 7, e7378 (2019).Article Google Scholar
- Stein, R. A., Sheldon, N. D. & Smith, S. Y. C3 plant carbon isotope discrimination does not respond to CO2 concentration on decadal to centennial timescales. N. Phytologist 229, 25762585 (2021).Article CAS Google Scholar
- Jardine, P. E. & Lomax, B. H. A 23 m.y. record of low atmospheric CO2. Geology 49, e523 (2021).Article ADS Google Scholar
- Franks, P. J. & Farquhar, G. D. The mechanical diversity of stomata and its significance in gas-exchange control. Plant Physiol. 143, 7887 (2007).Article CAS Google Scholar
- Kohn, M. J. Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proc. Natl Acad. Sci. 107, 1969119695 (2010).Article ADS CAS Google Scholar
- Fletcher, B. J., Brentnall, S. J., Quick, W. P. & Beerling, D. J. BRYOCARB: A process-based model of thallose liverwort carbon isotope fractionation in response to CO2, O2, light and temperature. Geochimica et. Cosmochimica Acta 70, 56765691 (2006).Article ADS CAS Google Scholar
- Porter, A. S., Yiotis, C., Montañez, I. P. & McElwain, J. C. Evolutionary differences in Δ13C detected between spore and seed bearing plants following exposure to a range of atmospheric O2:CO2 ratios; implications for paleoatmosphere reconstruction. Geochimica et. Cosmochimica Acta 213, 517533 (2017).Article ADS CAS Google Scholar
- Porter, A. S. et al. Testing the accuracy of new paleoatmospheric CO2 proxies based on plant stable carbon isotopic composition and stomatal traits in a range of simulated paleoatmospheric O2:CO2 ratios. Geochimica et. Cosmochimica Acta 2019, 6990 (2019).Article ADS Google Scholar
- Boyce, C. K. How green was Cooksonia? The importance of size in understanding the early evolution of physiology in the vascular plant lineage. Paleobiology 34, 179194 (2008).Article Google Scholar
- Streel, M. Late FrasnianFamennian climates based on palynomorph analyses and the question of the Late Devonian glaciations. Earth-Sci. Rev. 52, 121173 (2000).Article ADS Google Scholar
- Cleal, C. & Cascales-Miñana, B. Evolutionary Floras revealing large-scale patterns in Palaeozoic vegetation history. J. Palaeosci. 28, 120 (2021).
- Dahl, T. W. & Arens, S. K. M. The impacts of land plant evolution on Earths climate and oxygenation state An interdisciplinary review. Chem. Geol. 547, 119665 (2020).Article ADS CAS Google Scholar
- McMahon, W. J. & Davies, N. S. Evolution of alluvial mudrock forced by early land plants. Science 359, 10221024 (2018).Article ADS CAS Google Scholar
- Libertín, M., Kvaček, J., Bek, J., árský, V. & torch, P. Sporophytes of polysporangiate land plants from the early Silurian period may have been photosynthetically autonomous. Nat. Plants 4, 269271 (2018).Article Google Scholar
- Lenton, T. M., Daines, S. J. & Mills, B. J. W. COPSE reloaded: An improved model of biogeochemical cycling over Phanerozoic time. Earth-Sci. Rev. 178, 128 (2018).Article ADS CAS Google Scholar
- Krause, A. J. et al. Stepwise oxygenation of the Paleozoic atmosphere. Nat. Commun. 9, 4081 (2018).Article ADS Google Scholar
- Porada, P. et al. High potential for weathering and climate effects of non-vascular vegetation in the Late Ordovician. Nat. Commun. 7, 12113 (2016).Article ADS CAS Google Scholar
- Halder, S., Arens, S. K. M., Jensen, K., Dahl, T. W. & Porada, P. A dynamic local-scale vegetation model for lycopsids (LYCOm v1.0). Geosci. Model Dev. 15, 23252343 (2022).Article ADS CAS Google Scholar
- Bergman, N. M. COPSE: A new model of biogeochemical cycling over Phanerozoic time. Am. J. Sci. 304, 397437 (2004).Article ADS CAS Google Scholar
- McKenzie, N. R. et al. Continental arc volcanism as the principal driver of icehouse-greenhouse variability. Science 352, 444447 (2016).Article ADS CAS Google Scholar
- Glasspool, I. J., Edwards, D. & Axe, L. Charcoal in the Silurian as evidence for the earliest wildfire. Geol. 32, 381 (2004).Article ADS Google Scholar
- Glasspool, I. J. & Scott, A. C. Phanerozoic concentrations of atmospheric oxygen reconstructed from sedimentary charcoal. Nat. Geosci. 3, 627630 (2010).Article ADS CAS Google Scholar
- Sønderholm, F. & Bjerrum, C. J. Minimum levels of atmospheric oxygen from fossil tree roots imply new plant−oxygen feedback. Geobiology 19, 250260 (2021).Article Google Scholar
- Algeo, T. J., Scheckler, S. E. & Scott, A. C. Terrestrial-marine teleconnections in the devonian: links between the evolution of land plants, weathering processes, and marine anoxic events [and discussion]. Philos. Trans.: Biol. Sci. 353, 113130 (1998).Article Google Scholar
- Montañez, I. P. et al. Climate, pCO2 and terrestrial carbon cycle linkages during late Palaeozoic glacialinterglacial cycles. Nat. Geosci. 9, 824828 (2016).Article ADS Google Scholar
- Arora, V. K. & Montenegro, A. Small temperature benefits provided by realistic afforestation efforts. Nat. Geosci. 4, 514518 (2011).Article ADS CAS Google Scholar
- Grossman, E. L. & Joachimski, M. M. Oxygen Isotope Stratigraphy. in Geologic Time Scale 2020 279307 (Elsevier, 2020). https://doi.org/10.1016/B978-0-12-824360-2.00010-3.
- Chen, B. et al. Devonian paleoclimate and its drivers: A reassessment based on a new conodont δ18O record from South China. Earth-Sci. Rev. 222, 103814 (2021).Article CAS Google Scholar
- van Geldern, R. et al. Carbon, oxygen and strontium isotope records of Devonian brachiopod shell calcite. Palaeogeogr., Palaeoclimatol., Palaeoecol. 240, 4767 (2006).Article Google Scholar
- Liu, Y., Peltier, W. R., Yang, J. & Vettoretti, G. The initiation of Neoproterozoic snowball climates in CCSM3: the influence of paleocontinental configuration. Clim. Past 23, 25552577 (2013).
- Feulner, G. Climate simulations of Neoproterozoic snowball Earth events: Similar critical carbon dioxide levels for the Sturtian and Marinoan glaciations. Earth Planet. Sci. Lett. 404, 200205 (2014).Article ADS CAS Google Scholar
- Feulner, G. Formation of most of our coal brought Earth close to global glaciation. Proc. Natl. Acad. Sci. USA 114, 1133311337 (2017).Article ADS CAS Google Scholar
- De Vleeschouwer, D., Crucifix, M., Bounceur, N. & Claeys, P. The impact of astronomical forcing on the Late Devonian greenhouse climate. Glob. Planet. Change 120, 6580 (2014).Article ADS Google Scholar
- Brugger, J., Hofmann, M., Petri, S. & Feulner, G. On the sensitivity of the devonian climate to continental configuration, vegetation cover, orbital configuration, CO2 concentration, and insolation. Paleoceanogr. Paleoclimatology 34, 13751398 (2019).Article ADS Google Scholar
- Isbell, J. L. et al. Glacial paradoxes during the late Paleozoic ice age: Evaluating the equilibrium line altitude as a control on glaciation. Gondwana Res. 22, 119 (2012).Article ADS Google Scholar
- Gindre, L., Le Heron, D. & Bjørnseth, H. M. High resolution facies analysis and sequence stratigraphy of the Siluro-Devonian succession of Al Kufrah basin (SE Libya). J. Afr. Earth Sci. 76, 826 (2012).Article ADS Google Scholar
- Gerrienne, P., Bergamaschi, S., Pereira, E., Rodrigues, M.-A. C. & Steemans, P. An Early Devonian flora, including Cooksonia, from the Paraná Basin (Brazil). Rev. Palaeobot. Palynol. 116, 1938 (2001).Article Google Scholar
- Cookson, I. IVOn plant-remains from the Silurian of Victoria, Australia, that extend and connect floras hitherto described. Philos. Trans. R. Soc. Lond. B 225, 127148 (1935).Article ADS Google Scholar
- Earp, C. Costulatotheca schleigeri (Hyolitha: Orthothecida) from the Walhalla Group (Early Devonian) at Mount Pleasant, central Victoria, Australia. Alcheringa: Australas. J. Palaeontol. 43, 220227 (2019).Article Google Scholar
- Cohen, K. M., Finney, S. C., Gibbard, P. L. & Fan, J.-X. The ICS international chronostratigraphic chart. Episodes 36, 199204 (2013).Article Google Scholar
- Polissar, P. J., Fulton, J. M., Junium, C. K., Turich, C. C. & Freeman, K. H. Measurement of 13 C and 15 N Isotopic Composition on Nanomolar Quantities of C and N. Anal. Chem. 81, 755763 (2009).Article CAS Google Scholar
- Coplen, T. B. et al. New guidelines for δ 13 C measurements. Anal. Chem. 78, 24392441 (2006).Article CAS Google Scholar
- Bertrand, P. & Lu-Tao, W. Time-of-Flight Secondary Ion Mass Spectrometry (ToF-SIMS). in Microbeam and Nanobeam Analysis (eds. Benoit, D., Bresse, J.-F., Vant Dack, L., Werner, H. & Wernisch, J.) 167182 (Springer Vienna, 1996). https://doi.org/10.1007/978-3-7091-6555-3_8.
- Belu, A. M., Graham, D. J. & Castner, D. G. Time-of-flight secondary ion mass spectrometry: techniques and applications for the characterization of biomaterial surfaces. Biomaterials 24, 36353653 (2003).Article CAS Google Scholar
- Sodhi, R. N. S. Time-of-flight secondary ion mass spectrometry (TOF-SIMS):versatility in chemical and imaging surface analysis. Analyst 129, 483487 (2004).Article ADS CAS Google Scholar
- Pacanowski, R. & Griffies, S. The MOM 3 manual, alpha version. NOAA. Geophysical Fluid Dynamics Laboratory (1999).
- Hofmann, M. & M.A., Morales Maqueda. Performance of a second-order moments advection scheme in an Ocean General Circulation Model. J. Geophys. Res. 111, C05006 (2006).ADS Google Scholar
- Fichefet, T. & Morales Maqueda, M. A. Sensitivity of a global sea ice model to the treatment of ice thermodynamics and dynamics. J. Geophys. Res. 102, 1260912646 (1997).Article ADS Google Scholar
- Petoukhov, V. et al. CLIMBER-2: a climate system model of intermediate complexity. Part I: model description and performance for present climate. Clim. Dyn. 16, 117 (2000).Article Google Scholar
- Scotese, C. R. Atlas of Devonian Paleogeographic Maps, PALEOMAP Atlas for ArcGIS, The Late Paleozoic, Maps 65-72, Mollweide Projection. (2014).
- Belcher, C. M. & McElwain, J. C. Limits for combustion in low O2 redefine paleoatmospheric predictions for the mesozoic. Science 321, 11971200 (2008).Article ADS CAS Google Scholar